La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Pii: s0955-2863(99)00008-x
Feeding the nitric oxide synthase inhibitor L-Nnitroarginine elevates serum very low density lipoprotein and hepatic triglyceride synthesis in rats Tsuyoshi Goto,* Shoko Ohnomi,* Abdelkrim Khedara,* Norihisa Kato,* Hiroshi Ogawa,† and Teruyoshi Yanagita‡ *Department of Applied Biochemistry, Hiroshima University, Higashi-Hiroshima, Japan;†Department of Hygiene, Kinki University School of Medicine, Osaka, Japan; and ‡Department ofApplied Biological Sciences, Saga University, Saga, JapanThis study was conducted to study the influence of dietary L-Nnitroarginine (L-NNA), a nitric oxide (NO)synthase inhibitor, on serum lipids and lipoproteins and on the activities of enzymes related to lipid metabolismin rats. Feeding rats a diet containing 0.2 g/kg L-NNA for 5 weeks elevated serum concentrations of triglyceride,cholesterol, phospholipid, and free fatty acid and reduced serum nitrate (an oxidation product of NO). Theelevation in serum triglyceride was mainly due to the elevation in very low density lipoprotein (VLDL)triglyceride. Contents of cholesterol and phospholipid in the VLDL fraction also were elevated by L-NNA. L-NNAtreatment caused significantly higher activity of hepatic microsomal phosphatidate phosphohydrolase (therate-limiting enzyme in triglyceride synthesis) and lower activity of hepatic carnitine palmitoyltransferase (therate-limiting enzyme in fatty acid oxidation). Activities of hepatic enzymes responsible for fatty acid synthesissuch as glucose-6-phosphate dehydrogenase, malic enzyme, and fatty acid synthase were unaffected by L-NNA. The activity of hepatic microsomal phosphocholine cytidyltransferase (the rate-limiting enzyme in phosphatidyl-choline synthesis) was reduced significantly by L-NNA. Our results suggest that lower NO production caused theelevations in hepatic triglyceride synthesis by higher esterification of fatty acid and lower fatty acid oxidation,leading to an enrichment of VLDL triglyceride.
(J. Nutr. Biochem. 10:274 –278, 1999) Elsevier Science Inc.Keywords: nitric oxide; serum lipoproteins; hypertriglyceridemia; phosphatidate phosphohydrolase; carnitine palmitoyltransferase Introduction
which in turn causes some aggravation effects such ashypertension.9
Nitric oxide (NO) is an important cellular regulator.1,2 It has
Recently we have found that feeding L-Nnitroarginine
been shown to play roles in blood vessel dilation,1,2 immune
(L-NNA), which is a powerful specific inhibitor of NO
reactions,1,3 and the central and peripheral nervous sys-
synthase, to rats caused higher concentrations of serum
tems.1,2 NO production is enhanced by estrogen, inflamma-
triglyceride and cholesterol and lower serum nitrate (an
tion, and exercise through elevation of NO synthase activ-
oxidation product of NO).10 Adding excess L-arginine to
ity.4 – 8 NO is inactivated by reaction with superoxide
the diet containing L-NNA elevated serum nitrate by sup-
anion,1 and oxidative stress causes lower level of NO,
pressing competitive inhibition of NO synthase by L-NNA,and suppressed elevations of these lipids in serum. On thebasis of these facts, we speculate that lower NO productioncauses hyperlipidemia.10 Kurowska and Carrol11 also re-ported that feeding rabbits a diet containing the NO donor
Address correspondence to Dr. N. Kato, Department of Applied Biochem-
sodium nitroprusside caused a reduction in low density
istry, Hiroshima University, Higashi-Hiroshima, Japan. Received September 4, 1998; accepted January 29, 1999.
lipoprotein (LDL) cholesterol and a trend of reduction in
J. Nutr. Biochem. 10:274 –278, 1999 Elsevier Science Inc. 1999. All rights reserved.
655 Avenue of the Americas, New York, NY 10010
Higher VLDL by lower NO: Goto et al.
serum total cholesterol. Local generation of NO within the
Effect of dietary L-NNA on serum lipids and apolipoproteins
epicardial coronary arteries serves to inhibit platelet adhe-
sion and aggregation12 and to inhibit smooth muscle prolif-
eration.13 Therefore, lower NO generation seems to lead toatherosclerosis.
Our previous study provided evidence that hypercholes-
terolemia caused by L-NNA is mediated by lower synthesis
of bile acid from cholesterol,14 and that hypertriglyceride-
mia caused by L-NNA is due in part to lower hepatic fatty
acid oxidation.10 In this study, we further examined the
influence of L-NNA on serum lipoproteins and on hepatic
enzymes related to triglyceride synthesis in rats. Materials and methods
aSignificantly different from the contorl group (P Ͻ 0.05).
Male Wistar rats (Hiroshima Laboratory Animal Center, Hiro-
shima, Japan) weighing 50 to 70 g were used. Animals wereindividually housed in metal cages in a temperature-controlled(24°C) room with a 12-hour light-dark cycle (lights on, 8:00 am to
enzyme (ME), and fatty acid synthase (FAS) in the cytosol were
8:00 pm). All rats had free access to deionized water and experi-
assayed spectrophotometrically as described by Freedland21 and
mental diet. Composition of the basal diet was (in g/kg): casein,
Martin et al.,22 respectively. Activity of hepatic carnitine palmi-
200; sucrose, 217; ␣-corn starch, 433; corn oil, 50; cellulose
toyltransferase (CPT) in liver homogenate was measured using
powder, 50; salt mixture,15 35; vitamin mixture,15 10; DL-
L-carnitine, palmitoyl CoA, and 5,5Ј-dithio-bis (2-nitrobenzoic
methionine, 3; and choline bitartrate, 2. L-NNA (Aldrich Chemical
acid) according to the method of Bieber and Fiol.23
Company Inc., Milwaukee, WI USA) was added to the basal diet
Results were expressed as means Ϯ SE and analyzed by
at the level of 0.2 g/kg. After 5 weeks of consuming the diets, food
was removed from the cages at 8:00 am, and the rats were lightlyanesthetized with diethylether and euthanized between 1:00 pmand 3:00 pm. Blood was collected by heart puncture, and samples
were allowed to clot on ice. Serum samples were obtained bycentrifugation. Liver was immediately removed, weighed, and
Gain in body weight (g/5 wk) was unaffected by L-NNA
stored at Ϫ80°C until use. Portions of the fresh liver were used for
feeding (P Ͼ 0.05; control 285 Ϯ 4, L-NNA 270 Ϯ 6). Food
preparation of subcellular fractions.
intake (g/5 wk) also was unaffected by L-NNA (P Ͼ 0.05;control 712 Ϯ 14, L-NNA 685 Ϯ 18).
Serum concentrations of triglyceride, cholesterol, and
phospholipid were higher in the L-NNA group than in the
Serum lipoprotein fractions [very low density lipoprotein (VLDL),
control group (P Ͻ 0.05; Table 1). Serum free fatty acid was
d Ͻ 1.006 g/mL; LDL, d:1.006 –1.063 g/mL; and high densitylipoprotein (HDL), d:1.063–1.210 g/mL] were separated by step-
significantly elevated in the L-NNA group, whereas serum
wise density-gradient ultracentrifugation (TL-100, Beckman, San
ketone bodies were unaffected by L-NNA. Serum concen-
Francisco, CA USA).16 Total liver lipids were extracted by the
tration of nitrate was significantly reduced by L-NNA.
method of Folch et al.17 Concentrations of triglyceride, choles-
Serum concentrations of apo A-I and A-IV were signif-
terol, phospholipid, and free fatty acid were measured by kits from
icantly higher in the L-NNA group than the control group,
Wako Pure Chemical Co. (Osaka, Japan). Concentration of ketone
whereas concentrations of apo B and E were unaffected by
bodies (acetoacetate and 3-hydroxybutyrate) were measured by a
L-NNA. The ratio of apo B:apo A-I was unaffected by
kit (Ketone Test Sanwa Chemical Institute, Nagoya, Japan).
Concentrations of serum apolipoproteins (apo A-I, A-IV, B, and E)
Concentrations of triglyceride in the VLDL, LDL, and
were estimated by rocket electroimmunoassay.16 To estimate NO
HDL fractions were significantly higher in the L-NNA
production, serum concentration of nitrate (an oxidation product ofNO) was measured by a kit (Nitrate/Nitrite Assay Kit, Cayman
group than in the control group (P Ͻ 0.05; Figure 1).
Elevation in serum triglyceride by L-NNA treatment was
Portions of the fresh liver from individual rats were homoge-
due mainly to the elevation in VLDL triglyceride. L-NNA
nized in an ice-cooled 0.25 M sucrose solution containing a 10 mM
feeding also elevated VLDL cholesterol (P Ͻ 0.05),
Tris-HCl buffer (pH 7.4) and 1 mM EDTA. Microsomal and
whereas concentrations of cholesterol in the LDL and
cytosolic fractions were prepared as described previously.18 The
VLDL fractions were unaffected by L-NNA. Concentra-
fractions were stored at Ϫ80°C. Protein was assayed by a kit
tions of phospholipid in the VLDL and HDL fractions were
(Bio-Rad Protein Assay, Bio-Rad Laboratories, Richmond, CA
elevated by L-NNA (P Ͻ 0.05). Concentration of LDL
USA) using bovine serum albumin as the standard.
phospholipid was unaffected by L-NNA.
Activity of Mg2ϩ-dependent phosphatidate phosphohydrolase
Relative liver weight and concentrations of hepatic
(PAP) in liver microsomes and cytosol was assayed as describedpreviously.18 Activities of phosphocholine cytidyltransferase
cholesterol and phospholipid were unaffected by L-NNA
(CTP) in the microsomes and cytosol and of choline kinase (CK)
(Table 2; P Ͼ 0.05). There was a trend of elevation in liver
in the cytosol were measured by the reported methods.19,20
triglyceride concentration in rats that received L-NNA
Activities of glucose-6-phosphate dehydrogenase (G6PD), malic
(0.05 Ͻ P Ͻ 0.1). Activities of G6PD, ME, and FAS were
Distributions of triglyceride, cholesterol, and phospholipid among various lipoprotein fractions in rats fed diet with or without
L-Nnitroarginine (L-NNA). The vertical bars indicate the SE (n ϭ 10). *P Ͻ 0.05. VLDL, very low density lipoprotein; LDL, low density lipoprotein; HDL,high density lipoprotein.
unaffected by L-NNA. Activity of CPT, which is the
further demonstrated that the hypertriglyceridemia by L-
rate-limiting enzyme of mitochondrial -oxidation, was
NNA was ascribed mainly to a higher concentration of
reduced significantly by L-NNA addition.
triglyceride in VLDL fraction. Concentrations of choles-
The activity of microsomal Mg2ϩ-dependent PAP, which
terol and phospholipid in VLDL fraction also were clearly
controls the branching point in glycerolipid biosynthesis,
elevated by L-NNA, but not by very much. NO appears to
was elevated significantly by L-NNA, whereas the cytosolic
be an important regulator of serum VLDL triglyceride.
activity was unaffected (Table 3). Microsomal activity of
L-NNA treatment caused higher serum free fatty acid
CTP, the rate-limiting enzyme in phosphatidylcholine bio-
and lower activity of hepatic CPT (the rate-limiting enzyme
synthesis, was reduced significantly by L-NNA, whereas
of fatty acid oxidation) without affecting hepatic activities
the cytosolic activity was unaffected. Cytosolic activity of
of G6PD, ME, and FAS. This study further demonstrated
CK, the first enzyme on the de novo phosphatidylcholine
higher activity of PAP and lower activity of CTP in liver
biosynthesis pathway, was unaffected by L-NNA.
microsomes by L-NNA. It has been suggested that PAP andCTP are involved in the rate-limiting step of triglyceridesynthesis and phosphatidylcholine synthesis, respectively,
Discussion
and appear to exist in both soluble and particle forms, with
Consistent with our previous study10 was the finding that
the distribution of these forms being affected by the pre-
L-NNA treatment caused a marked hypertriglyceridemia.
vailing metabolic status.19,24 –26 The enzymes translocate
On the other hand, elevations in serum cholesterol and
from cytosol to the endoplasmic reticulum to become
phospholipid by L-NNA were only slight. The present study
functionally active and may help to regulate glycerolipidand phospholipid metabolisms.19,25 All of these resultssuggest that dietary L-NNA causes higher triglyceride
Effect of dietary L-NNA on liver lipids and the activities of
enzymes relating to fatty acid synthesis and oxidation in rats
Effect of dietary L-NNA on the activities of hepatic phos-
phatidate phosphohydrolase, phosphocholine cytidyltransferase, andcholine kinase in rats
aSignificantly different from the control group (P Ͻ 0.05). L-NNA–L-N nitroarginine. Higher VLDL by lower NO: Goto et al.
synthesis by increasing esterification of fatty acid and lower
Bredt, D.S. and Snyder, S.H. (1994). Nitric oxide: A physiologic
hepatic fatty acid oxidation, leading to the elevations of
messenger molecule. Ann. Rev. Biochem. 63, 175–195
Wink, D.A., Hanbauer, I., Krishna, M.C., DeGraff, W., Gamson, J.,
serum and VLDL triglyceride. On the other hand, the lower
and Mitchell, J. (1993). Nitric oxide protects against cellular damage
phosphatidylcholine synthesis also may indirectly enhance
and cytotoxicity from reactive oxygen species. Proc. Natl. Acad. Sci.
the synthesis of triglyceride by increasing fatty acids avail-
USA 90, 9813–9817
able for triglyceride synthesis. This assumption explains
Stadler, J., Stefanovic-Racic, M., Billiar, T.R., Curran, R.D., McIn-
why the elevation of serum triglyceride by L-NNA was
tyre, L.A., Greorgescu, H.I., Simmons, R.L., and Evans, C.H. (1991). Articular chondrocytes synthesize nitric oxide in response to
more prominent than that in serum phospholipid.
cytokines and lipopolysaccharide. J. Immunol. 147, 3915–3920
Recently we demonstrated that hypercholesterolemia
Beasley, D., Schwartz, J.H., and Brenner, B. M. (1991). Interleukin
caused by the inhibition of NO synthesis was at least in part
1 induces prolonged L-arginine-dependent cyclic guanosine mono-
mediated through lower activity of cholesterol 7 ␣-hydrox-
phosphate and nitrite production in rat vascular smooth muscle cells. J. Clin. Invest. 87, 602– 608
ylase, the rate-limiting enzyme of bile acid synthesis from
Bernstein, R.D., Ochoa, F.Y., Xu, X., Forfia, P., Shen, W. Thomp-
cholesterol.14 Activities of hepatic cholesterol 7 ␣-hydrox-
son, C.I., and Hintze, T.H. (1996). Function and production of nitric
ylase and CPT have been suggested to be regulated by
oxide in the coronary circulation of the conscious dog during
protein kinase C.27,28 This indicates that higher activity of
exercise. Circ. Res. 79, 840 – 848
protein kinase C may cause lower activities of both choles-
Koizumi, T., Hermo, C.I., Bjertnaes, L.J., Banerjee, M., Newman,J.H., and Kubo, K. (1996). Nitric oxide and nitroglycerin reversal of
terol 7 ␣-hydroxylase and CPT.27,28 It also has been
pulmonary vasoconstriction induced by alpha-activation during ex-
reported that acylation stimulating protein acts to stimulate
ercise. Am. J. Physiol. 270, H875–H880
triglyceride synthesis by increasing the rate of esterification
Hayashi, T., Yamada, K., Esaki, T., Kuzuya, M., Satake, S.,
of fatty acid via activation of protein kinase C.29 On the
Ishikawa, T., Hidaka, H., and Iguchi, A. (1995). Estrogen increasesendothelial nitric oxide by a receptor-mediated system. Biochim.
other hand, NO has been reported to inhibit the activity of
Biophys. Res. Commun. 214, 847– 855
protein kinase C.30 Therefore, we postulate that lower NO
Umans, J.G. and Levi, R. (1995). Nitric oxide in the regulation of
level might cause higher activity of hepatic protein kinase
blood flow and arterial pressure. Ann. Rev. Physiol. 57, 771–790
C, which in turn results in lower activities of cholesterol 7
Khedara, A., Kawai, Y., Kayashita, J., and Kato, N. (1996). Feeding
␣-hydroxylase and CPT and in higher activity of PAP,
rats the nitric oxide synthase inhibitor, L-Nnitroarginine, elevatesserum triglyceride and cholesterol and lowers hepatic fatty acid
leading to hyperlipidemia. Further study is in progress to
oxidation. J. Nutr. 126, 2563–2567
Kurowska, E.M. and Carrol, K.K. (1998). Hypercholesterolemic
Because apo B and E are main constituents of VLDL, we
properties of nitric oxide. In vivo and in vitro studies using nitric
expected that L-NNA treatment might elevate serum apo B
oxide donors. Biochim. Biophys. Acta 1392, 41–50
Azuma, H., Ishikawa, M., and Sekizaki, S. (1986). Endothelium-
and E. However, this possibility was eliminated by no
dependent inhibition of platelet aggregation. Br. J. Pharmacol. 88,
significant response of serum apo B and E to L-NNA
treatment. Synthesis and/or secretion of these apolipopro-
Sarkar, R. and Webb, R.C. (1998). Does nitric oxide regulate smooth
teins might not be involved in the effect of L-NNA on
muscle cell proliferation? A critical appraisal. J. Vasc. Res. 35,
Khedara, A., Goto, T., Kayashita, J., and Kato, N. (1998). Hyper-
Interestingly, L-NNA feeding resulted in significant
cholesterolemic effect in rats of a dietary addition of the nitric oxide
elevations in serum apo A-I and A-IV (Table 1). Higher
synthase inhibitor, L-Nnitroarginine, by less synthesis of bile acids.
levels of apo A-I and A-IV seem to relate to elevations in
Biosci. Biotechnol. Biochem. 62, 773–777
HDL triglyceride and phospholipid because these apoli-
American Institute of Nutrition (1977). Report of the AmericanInstitute of Nutrition and ad hoc committee on standards for
poproteins are major components of HDL. Recently, higher
nutritional studies. J. Nutr. 107, 1340 –1348
plasma apo A-IV level has been reported in hypertriglycer-
Ogawa, H., Sasagawa, S., Murakami, T., and Yoshizumi, H. (1995).
idemic patients.31 In addition, apo A-IV has been reported
Sesame lignans modulate cholesterol metabolism in the stroke-prone
to enhance the activation of lipoprotein lipase by apo C-II,
spontaneously hypertensive rat. Clin. Exp. Pharmacol. Physiol. 1,
which suggests a role in the metabolism of triglyceride-rich
Folch, J., Lees, M., and Sloane-Stanley, G.H. (1957). A simple
lipoproteins.32 Further study is in progress to examine the
method for the isolation and purification of total lipids from animal
influence of L-NNA treatment on the activity of lipoprotein
tissues. J. Biol. Chem. 226, 497–509
lipase or the utilization of serum triglyceride in adipose
Cha, J.-T., Maeda, Y., Yamamoto, K., Oogami, K., and Yanagita, T.
(1998). Association between hepatic triacylglycerol accumulationinduced by administering orotic acid and enhanced phosphatidate
A recent study by Minami et al.33 demonstrated that
phosphohydrolase activity in rats. Biosci. Biotechnol. Biochem. 62,
triglyceride-rich human plasma suppressed the NO synthe-
sis in human endothelial cells. Together with our study,
Wright, P.S., Morand, J.N., and Kent, C. (1985). Regulation of
lower NO appears to induce hypertriglyceridemia, which in
phosphatidylcholine biosynthesis in Chinese hamster ovary cells byreversible membrane association of CTP: Phosphocholine cytidylyl-
turn causes further suppression of NO production, leading to
transferase. J. Biol. Chem. 269, 7919 –7926
exaggeration of some aggravation effects such as hyperten-
Ishidate, K. Iida, K., Tadokoro, K., and Nakazawa, Y. (1985).
Evidence for the existence of multiple forms of choline (ethanol- amine) kinase in rat tissues. Biochim. Biophys. Acta 833, 1– 8
Freedland, R.A. (1967). Effect of progressive starvation on rat liver enzyme activities. J. Nutr. 91, 489 – 495 References
Martin, D.B., Horning, M.G., and Vagelos, P.R. (1961). Fatty acidsynthesis in adipose tissue, I. Purification and properties of a long
Moncada, S., Palmer, R.M., and Higgs, E.A. (1991). Nitric oxide:
chain fatty acid-synthesizing system. J. Biol. Chem. 236, 663– 668
Physiology, pathophysiology and pharmacology. Pharmacol. Rev.
Bieber, L.L. and Fiol, C. (1985). Purification and assay of carnitine
43, 109 –142
acyltransferases. Methods in Enzymol. 12, 276 –284
Lamb, R.G. and Fallon, H.J. (1974). Glycerolipid formation from
flone, K.K. (1995). Single transduction pathway of acylation stim-
sn-glycerol-3-phosphate by rat liver cell fractions. The role of phos-
ulating protein: Involvement of protein kinase C. J. Lipid Res. 36,
phatidate phosphohydrolase. Biochim. Biophys. Acta 348, 166 –178
Fremont, L. and Gozzelino, M.T. (1996). Dietary sunflower oil
Gopalakrishna, R., Chen, Z.H., and Gundimela, U. (1993). Nitric
reduces plasma and liver triacylglycerols in fasting rats and is
oxide and nitric oxide-generating agents induce a reversible inacti-
associated with decreased liver microsomal phosphatidate phospho-
vation of protein kinase C activity and phorbol ester binding. J. Biol.
hydrolase activity. Lipids 31, 871– 878 Chem. 268, 27180 –27185
Jamal, Z., Martin, A., Gomez-Munosz, A., and Brindley, D.N.
Verges, B., Rader, D., Schaefer, J., Zech, L., Kindt, M., Fairwell, T.,
(1991). Plasma membrane fractions from rat liver contain a phos-
Gambert, P., and Brewer, H.B., Jr. (1994). In vivo metabolism of
phatidate phosphohydrolase distinct from that in the endoplasmic
apolipoprotein A-IV in severe hypertriglyceridemia: A combined
reticulum and cytosol. J. Biol. Chem. 266, 2988 –2996
radiotracer and stable isotope kinetic study. J. Lipid Res. 35,
Stravitz, R.T., Vlahcevic, Z.R., Gurley, E.C., and Hylemon, P.B.
(1995). Repression of cholesterol 7 ␣-hydroxylase transcription by
Goldberg, I.J., Scheraldi, C.A., Yacoub, L.X., Saxena, U., and
bile acids is mediated through protein kinase C in primary cultures of
Bisgaier, C.L. (1990). Lipoprotein ApoC-II activation of lipoprotein
rat hepatocytes. J. Lipid Res. 36, 1359 –1369
lipase. Modulation by apolipoprotein A-IV. J. Biol. Chem. 265,
Harano, Y., Kashiwagi, A., Kojima, H., Suzuki, M., Hashimoto, T.,
and Shigeta, Y. (1985). Phosphorylation of carnitine palmitoyltrans-
Minami, M., Yokokawa, K., Kohno, M., Yasunari, K., Hanehira, T.,
ferase and activation by glucagon in isolated rat hepatocytes. FEBS
and Yoshikawa, J. (1997). Suppression of histamine-induced nitric
Lett. 188, 267–272
oxide synthesis by triglyceride in human endothelial cells. Jap.
Baldo, A., Sniderman, A.D., St. Luce, S., Zhang, X.-J., and Cian-
J. Pharmacol. 75(suppl. I), 55P
SCHEME FOR MOBILE LEGAL SERVICES-CUM- LOK ADALAT IN MAHARASHTRA v OBJECT : The Legal Aid is an essential part of the administration of justice. The goal isto secure justice for the weaker and downtrodden sections of the Society. The de-mocracy has no meaning for the citizens unless they are able to secure their basicrights namely Education, Employment and right to live life of dignity
DREIFALTIGKEITS-KRANKENHAUS WESSELINGAbteilung für Plastische Chirurgie Chefarzt: Dr. med. Dirk F. Richter Anmelde- / Anamnesebogen Name / Vorname:_____________________________________________________________Straße: _____________________________________________________________________PLZ: ____________ Ort: ______________________________________________________ Telefon (Privat) : ________
 Feeding the nitric oxide synthase
Feeding the nitric oxide synthase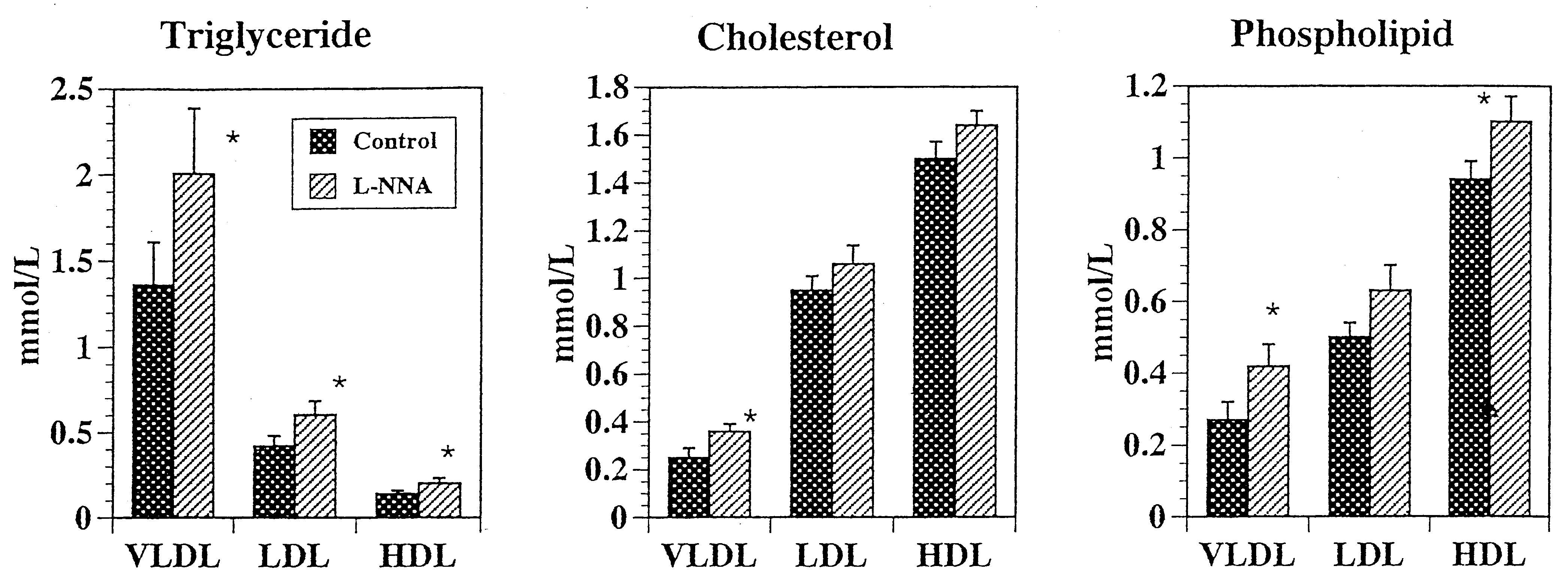 Distributions of triglyceride, cholesterol, and phospholipid among various lipoprotein fractions in rats fed diet with or without
L-Nnitroarginine (L-NNA). The vertical bars indicate the SE (n ϭ 10). *P Ͻ 0.05. VLDL, very low density lipoprotein; LDL, low density lipoprotein; HDL,high density lipoprotein.
Distributions of triglyceride, cholesterol, and phospholipid among various lipoprotein fractions in rats fed diet with or without
L-Nnitroarginine (L-NNA). The vertical bars indicate the SE (n ϭ 10). *P Ͻ 0.05. VLDL, very low density lipoprotein; LDL, low density lipoprotein; HDL,high density lipoprotein.