La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Dspace.cuny.edu
Chronic Food-Restriction Decreases Brain Serotonin Transporter Sites in the Rat Richard Paternoster
Submitted to the Committee on Undergraduate Honors at Baruch College of the City University of New York in partial fulfillment of the requirements for the degree of Bachelor of Arts in Biology with Honors.
List of AbbreviationsSummaryAbstractIntroductionMaterials and MethodsResultsDiscussionAcknowledgementsReferencesFigures and Table 1
LIST OF ABBREVIATIONS ACTH- Adrenocorticotropic hormone Cl, C2, C3, C4- Control Group (Ad Libitum) Rats CRH- Corticosterone-releasing hormone fmols- femtomoles ( 1 x 10-15 moles) FR1, FR2, FR3, FR4- Food-Restricted Rats g- grams HPA- Hypothalamic-pituitary-adrenal axis LH- Lateral Hypothalamus Mg- milligrams ml- milliliters mM- millimolar (1 x 10-6 Molar) mRNA- Messenger RNA nm- nanometer (1 x 10-9 meters) nM- nanomolar (1 x 10-9 Molar) 3H-paroxetine- Tritiated (radioa ctively labeled) paroxetine PCPA- p-chlorophenylalanine pmols- picomoles (1 x 10-12 moles) PVN- Paraventricular nucleus of the hypothalamus RPM- Revolutions per minute TrOH- Tryptophan hydroxylase enzyme ul- microliter (1 x 10-6 liters) uM- micromolar (1 x 10-6 Molar) 5-HT- 5-hydroxytryptamine (Serotonin) 5-HTP- 5-hydroxytryptophan (Serotonin metabolite)
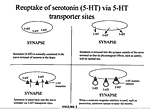
Serotonin (5-HT), a chemical found in various brain regions including the hippocampus, brainstem, cortex, and striarum, acts as a neurotransmitter, i.e., it transmits messages from one nerve cell to another. Certain nerve cells release 5-HT into the synapse, the space between two or more communicating nerve cells. Once in the synapse, 5-HT influences the behavior of nearby nerve cells by acting on specific 5-HT receptors on their surfaces. The effects of 5-HT are terminated by the removal of 5-HT from the synapse by the same nerve cells that originally released it. The reuptake of 5-HT by these cells is carried out by specific 5-HT transporter sites in their cell membranes.
Disturbances in neurotransmission mediated by 5-HT may have a role in depression, anxiety and other psychological disorders. Serotonin's most studied effect is that on appetite. Experimentally elevated brain 5-HT levels are correlated with an activation of satiety and decreased food ingestion in rats. Food-restriction is a widely used protocol for obtaining decreased 5-HT levels in rats. Decreased brain 5-HT levels can also be obtained by treating animals with p-chlorophenylalanine (PCPA), an inhibitor of 5-HT synthesis.
The present study examines the relationship between chronic food-restriction and 5-HT transporter site concentration in rats. Chemical depletion of 5-HT by treatment with PCPA has been shown to decrease the concentration of 5-HT transporter site messenger RNA (mRNA), a molecule required for the synthesis of new 5-HT transporter sites. A decrease in 5-HT transporter site mRNA concentration would. be expected to lead to a decrease in 5-HT transporter sites.
Because no one has yet studied the relationship between food-restriction and 5-HT transporter site density in rats, we decided to determine if chronic food-restriction causes a decrease in the 5-HT transporter site density. Knowledge of this decrease in rats due to food-restriction may be used as an animal model to better study bulimia, obesity, and depression, which are treated effectively with 5-HT reuptake inhibitors such as Prozac, fluoxetine, and sertraline.
Experimentation was carried out utilizing a total of 24 male Sprague-Dawley rats. These 24 rats were separated into 3 experiments of 8 rats each. The three experiments were carried out at different periods. In each group, 4 rats were kept on a food-restricted diet of
10g of food per day and the remaining 4 rats were fed ad libitum (intake is approximately 20-30g of food per day) for 14 days. Therefore, 12 rats in total were food-restricted and 12 were fed ad libitum. At the end of the 14 days, all of the subjects were sacrificed. The hippocampus, striaturn, brainstem, and cortex regions of the brain were removed, frozen, and stored. The density of 5-HT transporter sites (the sites at which released 5-HT is taken back into the cell) was estimated by measuring paroxetine-binding by membranes from cells in these brain regions. Paroxetine is known to bind to 5-HT transporter sites.
In this study, food-restriction produced a significant decrease in 5-HT transporter sites in the brainstem and hippocampus in two of the three experimental groups. The level of 5-HT transporter sites was significantly decreased in the striatum in only one of the experimental groups. Although a sizable decrease (34%) was seen in the striatum of a second group, the finding was not significant due to a large standard deviation. Food-restriction produced no decrease in 5-HT transporter sites in the cortex for any experimental group.
The results from one of the experimental groups (group 2) differed from the others. The levels of 5-HT transporter sites were higher in the group 2 rats (in both control and food-restricted animals) than in the other groups, and the decrease in 5-HT transporter sites seen in food-restricted rats in groups I and 3 was not found in the group 2 rats. The group 2 rats were from a different commercial supplier. Perhaps the group 2 rats were slightly different genetically or perhaps they were fed or handled differently prior to their arrival at the laboratory. These hypotheses remain to be tested.
In summary, we obtained positive results from the 5-HT transporter site density assays, showing that food-restriction not only decreases the concentration of brain 5-HT, but also decreases the 5-HT transporter site density. Since there is a decrease in the concentration of 5-HT in the synapse during periods of chronic food-restriction, the cell may compensate for this decrease by keeping as much of this reduced amount of 5-HT as possible in the synapse. This may be achieved by reducing the amount of 5-HT transporter sites, through which 5-HT is taken back into the cell. This information could be very helpful in future endeavors to treat appetite disorders such as bulimia and obesity and psychological disorders such as depression in which 5-HT has a putative role. ABSTRACT
Chemical depletion of brain serotonin (5-HT) by the synthesis inhibitor, p-chlorophenylalanine, decreases 5-HT transporter mRNA levels, displaying a positive relationship between the levels of brain 5-HT and 5-HT transporters. Since food-restriction is known to cause a decrease in serotonin levels in certain brain regions, a concomitant decrease in 5-HT transporter site density due to food restriction might be expected. In this study, selected brain regions of rats on food-restricted and ad libitum diets were dissected and the density of 5-HT transporter sites in these regions was measured using paroxetine-binding assays. It was found that food-restricted rats had
decreased levels of 5-HT transporter sites in the hippocampus, brainstem, and striatum. Such information could be useful in an animal model in the study of bulimia, obesity, and depression, disorders which are effectively treated with 5-HT reuptake inhibitors. INTRODUCTION
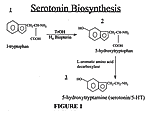
Serotonin, 5-hydroxytryptamine (5-HT), is a neurotransmitter whose synthesis begins with the binding of the amino acid L-tryptophan to the enzyme tryptophan hydroxylase (TrOH)- the rate limiting enzyme in the synthesis of 5-HT. The addition of oxygen and the cofactor H4 Biopterine allows for the hydroxylation of L-tryptophan thereby
producing 5-hydroxytryptophan (5-HTP). A cofactor is a substance, usually a metal, which is frequently required by an enzyme to catalyze a reaction. Hydroxylation is the addition of a hydroxyl group (-OH) to a molecule. In this case, L-tryptophan is hydroxylated to produce 5-HTP. An L-amino acid decarboxylase enzyme then removes a carboxyl group (-COOH) from 5-HTP to produce serotonin (2) (Figure 1).
The rate-limiting step in the synthesis of 5-HT is the activity of TrOH (2) and is probably directly correlated with the available amount of L-tryptophan which comes from the diet (37). The Km value of L-tryptophan, which represents the concentration ofsubstrate which
allows the enzyme to function at half of its maximum rate, for TrOH is approximately 50 uM (0.05 mM) (12).
5-HT axons are located in many regions of the brain. The most densely 5-HT-innervated regions in rats are the frontal cortex, amygdala, basal forebrain, and most of the brainstem. The posterior cerebral cortex, parts of the diencephalon, cerebellum and white matter are less densely innervated (36). In the forebrain, 5-HT-containing fibers emanate from two nuclei in the brainstem. Fibers from the dorsal raphe nuclei contain axon terminals with fine varicosities (approx. 1-2 um). Fibers from the median raphe nuclei contain axon terminals with larger varicosities (approx. 3-4 um) (25,28). Reductions in 5-HT levels in the forebrain of both rodents and non-human primates have been shown to occur after administration of several amphetamines, namely 3,4-methylenedioxyamphetamine (33), 3,4-methylenedioxymethamphetamine (12), and N-ethyl-3,4-methylenedioxyamphetamine (34). These amphetamines are used recreationally by humans and denervate the fine-fibered axons (36). Although 5-HT axons are found throughout the brain, the largest reservoirs of 5-HT are located in the blood and in the gastrointestinal mucosa (20).
5-HT axons also display a degree of regeneration. Ten days subsequent to the injection of a neurotoxin that causes the destruction of 5-HT containing axons (2), into the spinal cord of animals whose axons had been damaged, it was found that new axons had appeared in the medulla oblongata (2). After three months, new fibers were also seen in the spinal cord. Normal connections and projections were noted along with abnormal ones (2).
5-HT is thought to have a modulatory role on temperature, pain, sex behavior, the regulation of sleep, locomotor activity, avoidance learning, water consumption, self-stimulation, and aggression (2). 5-HT may also play a role in psychiatric disorders. Impulsivity, depression, anxiety and obsessive-compulsive disorders may be caused, in part, by disturbances in the neurotransmission mediated by 5-HT (10,40). 5-HT is also thought to have a role in the onset of migraines (18), cerebral vasospasms, and in cerebral blood flow (9).
One of the most studied roles of 5-HT deals with appetite. Elevated 5-HT levels have been shown to cause the activation of satiety in food-restriction studies (6,7,13). These studies have utilized 5-HT reuptake inhibitors, e.g., fenfluramine and fluoxetine, and 5-HT receptor antagonists, e.g., metergoline and methysergide. Reuptake inhibitors prevent 5-HT from binding to its transporter and result in a greater amount of extracellular 5-HT because of a decreased amount of 5-HT allowed back into the cell. Antagonists are drugs which block the receptors of a neurotransmitter and therefore inhibit the neurotransmitter from binding to the receptor. Fenfluramine, which is most effective in the paraventricular nucleus (PVN) of the hypothalamus, has an anorectic effect on rats (41), and fluoxetine causes a decrease in the size of the meals consumed by food-restricted rats (11,25,27). This decrease in the size of meals consumed is blocked by the administration of metergoline, a 5-HT2A receptor antagonist (21). Methysergide, a second 5-HT2 receptor
antagonist, causes a stimulation of feeding in rats which are well-sated (25,39). Furthermore, food intake inhibition has also been observed after administration of 5-HT1B
receptor agonists (22-24,27,38). This information displays that appetite is reduced by elevated levels of free 5-HT.
5-HT receptors are classified according to three definitive criteria: operational (the drug-related characteristics of the receptor), structural, and transductional (receptor-effector coupling events). These transductions occur via a second messenger system whereby the binding of a chemical to a receptor leads to a change in the concentration of the second messenger, e.g., adenyl cyclase. This change in concentration usually leads to an alteration in the activity of an enzyme and therefore results in the eventual physiological effect of the chemical. 5-HT2 receptors are structurally different from 5-HT1 receptors,
they are affected by different drugs, and transduce their effects through phosphoinositide metabolism as compared to 5-HT1 receptors which transduce their effects by being
negatively linked to adenyl cyclase (17).
The effects of 5-HT on appetite are not only seen through the use of antagonists, agonists, and reuptake inhibitors, but also directly with 5-HT. Administration of 5-HT into the PVN was shown to reduce carbohydrate intake by rodents (26). Carbohydrate intake wasalso attenuated when 5-HT was administered into the medial hypothalamus of rodents given a high-protein and high-carbohydrate diet (39). Therefore, increased extracellular 5-HT causes reduced carbohydrate intake.
This effect of 5-HT may be explained through an influence of 5-HT on the lateral hypothalamus (LH). Neurons in the LH initiate feeding behavior and 5-HT has been shown to have an inhibitory effect on these neurons (42). Rodents do not have the ability
to vomit. Instead, when an unfavorable substance has been ingested, a conditioned taste aversion ensues where the rodent will no longer ingest that substance. A conditioned taste aversion to saccharin solution can be instilled in rodents. When a saccharin solution, to which a conditioned taste aversion had been instilled, was orally infused into rodents, elevated levels of 5-HT were observed in the LH. The same elevated levels were also noted in rodents receiving injections of lithium chloride (LiCl), which is illness inducing. (42).
Chemical depletion of brain 5-HT due to administration of the 5-HT synthesis inhibitor p-chlorophenylalanine (PCPA) has been shown to be correlated with a decrease in 5-HT transporter mRNA levels (32). 5-HT levels are elevated during and after a meal and are decreased during periods of food-restriction (29). Since a positive correlation has been observed between the concentration of brain 5-HT and mRNA levels of 5-HT transporters, 5-HT transporter site density may also be decreased during periods of food-restriction. In this study, we desired to determine if, in fact, chronic food-restriction does cause a decrease in 5-HT transporter site density. MATERIALS AND METHODS Food-Restriction
Twenty-four male Sprague-Dawley rats weighing from 300-350g were separated into three experimental groups of eight rats each. These eight rats were individually caged and used during one two-week period of food-restriction. Each group of rats was subjected to the following protocol at different periods. They arrived at the NYU animal facility on Day I and were given three days to become accustomed to the surroundings prior to food-restriction. On Day 4 food carriers were removed from the cages of two of the food-restricted subjects. Food carriers with unlimited food were retained in the cages of control, ad libitum, rats. These two subjects were designated FR1 and FR2, and two control rats were designated Cl and C2. On Day 5, the food carriers were removed from the cages of the remaining two food-restricted subjects. These subjects were designated FR3 and FR4 and the final two rats were designated C3 and C4. Also on Day 5, subjects FR1 and FR2 were given only 10g of food pellets between the hours of 2PM and 5PM and were fed in this manner for the duration of the experiment. Subjects FR3 and FR4 were started on their 10g of food pellets per day diet on Day 6. Controls were fed ad libitum and their typical daily intake was in the range of 20 to 30 grams.
This staggered feeding schedule was utilized in order to sacrifice only four subjects per day after the 14 days of food-restriction. On Day 18, subjects C1, C2, FR1, and FR2 were sacrificed approximately at 2PM. On this day, no food was given to subjects FR1 and FR2. Subjects FR3 and FR4 were sacrificed on Day 19.
The method used to sacrifice the subjects was to first anesthetize them with carbon dioxide and then decapitate them with a guillotine. Several regions of the brain-
hippocampus, cortex, brainstem, and striatum- were then dissected and placed on dry ice. These regions were stored at -70 C until further use. The hippocampus, cortex, brainstem, and striatum were utilized because they are regions which contain the greatest amount of serotonergic innervation. Homogenization of Brain Regions
The dissected sections of the subject's brains - brainstem, cortex, hippocampus and striatum - were removed from the -70 C freezer and individually weighed. Each section was then placed into a separate centrifuge tube containing 7 ml of paroxetine buffer (composed of 38.5 mM Tris-HCl, 11.5mM Tris-Base, 120mM NaCl, and 5mM KCl), pH 7.4, and homogenized for 10 second's utilizing a Polytron homogenizer. Following homogenization, the samples were centrifuged for 20 minutes at 17,000 RPM to isolate plasma membrane fractions. Subsequently, the supernatant was removed and the pellet re-suspended in a 1:80 dilution of paroxetine buffer. The weight of each brain region was multiplied by 80 and that value became the amount of paroxetine buffer added to each tube. Samples were once again homogenized, placed into separate 50 ml plastic conical tubes, and then frozen at -70 C until use. Prior to freezing, however, I ml of each homogenate was pipetted into 1 ml microfuge tubes and frozen separately for use in a Lowry protein assay. Lowry Protein Assay
The homogenized tissue samples stored at -70 C were brought to room temperature. 160 ul of the tissue was pipetted into separate glass test tubes followed by 840 ul of double-distilled H20. Then, 1 ml of ABC solution (0.15% Sodium deoxycholate; 72%
trichloroacetic acid; copper tartrate/carbonate solution; 20% Folin-Ciocalteu reagent) was added to each tube, and the tubes were vortexed. After 15 minutes of incubation at room temperature, 3 ml of Folin's Reagent (2N Phenol Reagent Solution), in a 1:10 dilution, was added to each tube, the tubes were vortexed, and incubation at room temperature was continued for 1 hour. 1 ml of the mixture was then removed from the tubes and placed into cuvette for spectrophotometrical analysis at a wavelength of 570 nm.
Standards were prepared utilizing concentrations of bovine serum albumin ranging from 100 ug/ml to 1 mg/ml. The results were interpreted and converted to mg/ml of protein utilizing the Radlig EDBA Version 3.0 program by McPherson. These results were later used to calculate the fmol of bound paroxetine per mg of protein in the sample. 3H-Paroxetine Binding Assay
The frozen, homogenized tissue samples were brought to room temperature and 160 ul of each was added to eight plastic test tubes. Four of these tubes contained only 40 ul of paroxetine (0.25 nM) without any fluoxetine to test for total binding, and the remaining four contained 40 ul of fluoxetine (1 uM) to obtain the amount of non-specific binding. Fluoxetine blankets the binding sites and therefore displaces the 3H-paroxetine. This allowed for the detection of non-specific binding, which was subtracted from the total
binding to obtain the specific binding value of 3H-paroxetine. These tubes were then vortexed and incubated for 15 minutes at room temperature. Subsequently, 200 ul of 3H-paroxetine buffer was added to each tube. The samples were then incubated for 120 minutes at room temperature. Following this final incubation, the samples were filtered over 0.1% polyethyleneimine (PEI) coated filters utilizing a Brandell cell harvester. PEI prevents any free ligand from binding to the filter. Each glass fiber filter was then placed into a glass test tube which was filled with 5 ml of Ecoscint A scintillation fluid. The tubes were then placed into a scintillation counter to count the amount of 3H-paroxetine bound to the rat brain membrane homogenates.
The values obtained for the fluoxetine binding of each tissue sample, i.e., Subject C1 Cortex, Subject C1 Brainstem, etc., were subtracted from those obtained from the paroxetine binding of the same sample. This new value was the amount (pmol) of specific binding of 3H-paroxetine to serotonergic transporter sites. This specific binding value was divided by the specific activity of the 3H-paroxetine, (21 Curies/mmole) 24,000 counts per minute/pmol, to obtain the amount of paroxetine bound to the terminal preparations (membrane homogenates). Finally, this amount of bound paroxetine was divided by the concentration of protein of each sample obtained from the Lowry assay to result in the fmols of paroxetine bound to terminal preparations/mg of protein. Statistical analysis was conducted utilizing a two-tailed Student's t-test. All data points were recorded as Mean + SEM of four animals per group. Weight Changes Caused by Food Restriction
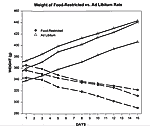
On the day of sacrifice, food-restricted rats weighed approximately 85% of their initial starting weight, whereas ad libitum rats weighed approximately 120% of their initial starting weight. Mean body weights and percentages of starting weights are listed in Table 1 and plotted in Figure 3. Quantification of 5-HT Transporter Sites Hippocampus
In two of the three paroxetine-binding assays (experimental groups 1 and 3), there were significant reductions in the density of 5-HT transporter sites in the hippocampus of food-restricted rats (Figure 4). In the first experimental group, there was a decrease of 39% in the density of 5-HT transporter sites in the hippocampus in the food-restricted subjects as compared to the control subjects. Specifically, the control subjects had an average of 156 fmol/mg of specific 3H-paroxetine binding compared to 96 fmol/mg of specific 3H-paroxetine binding in food-restricted subjects. The p values, which denote the level of significance chosen as our standard, were required to be below 0.05 in all of the assays. The p value for this experimental group was 0.003. The third experimental group also
displayed a decrease in the 5-HT transporter sites in food-restricted subjects. The difference was 21% between food-restricted and control subjects, and specifically there was 262 fmol/mg average specific 3H-paroxetine binding in the control subjects compared to 207 fmol/mg in the food-restricted subjects. The p value for the third experimental group was 0.01.
The second experimental group did not display any siginificant decrease in 5-HT transporter sites. The p value here was 0.54. Brainstem
As in the hippocampus, significant reductions in 5-HT transporter site density was also seen in the breinstem in experimental groups 1 and 3 (Figure 5). The reduction from control to food-restricted subjects was 19% in the first experimental group. The values were 187 fmol/mg of specific 3H-paroxetine binding for the controls compared to 151 fmol/mg for the food-restricted subjects. The p value obtained was 0.01. This reduction was approximately 20% from control to food-restricted subjects in the third experimental group, and the values were 179 fmol/mg of specific 3H-paroxetine binding forthe controls compared to 143 fmol/mg for the food-restricted subjects. The p value was 0.001.
Also as for the hippocampus, the second experimental group showed no significant decrease in 5-HT transporter site decrease. The p value for the second experimental group was 0.26.
The cortex displayed no significant reduction in any of the three experimental groups as can be seen in Figure 6. The p values for the cortex of all three experimental groups are 0.41, 0.81, and 0.185. Striarum
A significant reduction in 5-HT transporter sites in the striatum was shown in only one of the three experimental groups (Figure 7). In experimental group 3, there was a reduction of 27.6% in the 5-HT transporter site density from control subjects to food-restricted subjects. The p value for this experimental group was 0.01.
Reduction from control subjects to food-restricted subjects was 34% in the first experimental group, but the p value for this group was 0.239, due to a high value for the standard deviation. Therefore, there was no significant difference in 5-HT transporter site density in this group.
As for the other three brain regions, the second experimental group displayed no significant reduction in 5-HT transporter site density. The p value was 0.29. DISCUSSION
In the present study, food-restriction appears to lower 5-HT transporter site density in some brain regions. In two of the three experimental groups, food-restriction was shown to cause a significant decrease in 5-HT transporter sites in the hippocampus and brainstem, and in the striatum in one of the three experimental groups. A large percent reduction (39%) in striatal 5-HT transporter site density was seen in a second of the three experimental groups, however a large value for the standard deviation (120.9) denoted that a wide range of 3H-paroxetine binding values of the control subjects existed, and therefore the results were not significant. The results in the second experimental group differed markedly from the other two groups. No differences in the density of 5-HT transporter sites between ad libitum and food-restricted rats were observed in any brain region in the second group of rats.
The only known variable in the second experimental group which was not seen in the other two groups was that the rats were obtained from a different supplier. A different manner in which they were bred by the supplier, e.g., diet, or a genetic variation due to the particular breeding of the rats may have led to the difference in the binding of paroxetine to the 5-HT transporter. This hypothesis is not proven, but it would be interesting to observe if the same high value of 5-HT transporter site density was againobserved under the identical circumstances. If, in fact, this was the case, it would also be interesting and beneficial to see if a difference in the breeding of the rats does take place, and if the binding of paroxetine is effected by this factor.
Food-restriction, as presently reported, does result in a decrease in the amount of 5-HT in certain brain regions. Less 5-HT is produced by the neurons and therefore, extracellularly there is probably less 5-HT. A reason for the decreased level of transporters may be that in order to compensate for this reduced level extracellularly, fewer transporters may exist to ensure that a reduced amount of 5-HT is taken back into the neurons. Immunocytochemical analysis may be helpful to discover the amount of tryptophan hydroxylase present in certain brain regions and possibly provide evidence showing whether or not food-restriction may have a harmful effect in the synthesis of 5-HT.
The evidence obtained in this study that 5-HT transporter site density in rats is reduced in certain brain regions due to chronic food-restriction may contribute to our understanding of bulimia and other eating disorders. Bulimia is clinically treated with 5-HT reuptake inhibitors such as fluoxetine which binds to the transporter sites on the nerve terminal and prevents 5-HT from re-entering the cell. There seems to be, then, a significant correlation between 5-HT transporter sites and the effective treatment of bulimia. This current data may be used as an animal model in the study of bulimia.
Results of this study suggest further lines of investigation. There is various evidence which points to the existence of a strong correlation between the serotonin system and the glucocorticoid system. Glucocorticoids are steroids which promote gluconeogenesis (29). They are produced and secreted via the hypothalamic-pituitary-adrenal (HPA) axis (Figure 8). This axis is initiated by the release of corticosterone-releasing hormone
(CRH) from the hypothalamus to induce the basophilic cells of the anterior pituitary to produce, as well as release, adrenocorticotropic hormone (ACTH). Several neurotransmitters, one of which is serotonin, indirectly stimulate the release of CRH at this point of the cycle via 5-HT1A and 5-HT2A/2C receptors (29). Glucocorticoids affect the
serotonin system by stimulating 5-HT synthesis and stress-induced tryptophan hydroxylase activity (3,4,5). Further correlation is observed through the fact that glucocorticoids are also affected by food-restriction. Plasma corticosterone (the major glucocorticoid in rats) levels have been shown to be elevated preceding the scheduled daily meal in food-restricted rats (1). Since abnormalities in the HPA axis have been shown to be correlated to depression (35,45) and one of the major aberrant behavioral effects of disturbances in the neurotransmission of 5-HT is depression (10,40), experimentation regarding the correlation between the effects of food-restriction on both systems could prove useful. ACKNOWLEDGEMENTS
I would like to extend my appreciation to Dr. Eric J. Simon for his generous support of this study, and to Drs. H. Kenneth Kramer, Glen Abrahamsen, and Kenneth Carr for their interest and invaluable assistance. I would also like to thank the Baruch College Natural Sciences Departmental Honors Committee for their continuing support and advice. REFERENCES
1. Abrahamsen, G.C.; Berman, Y.; Carr, K.D. Curve-shift analysis of self-stimulation in food-restricted rats: relationship between daily meal, plasma corticosterone, and reward sensitization. Brain Research 695:186-194; 1995.
2. Azmitia, E.C.; Conrad, L.C. Temporal effects of formix transection on brain tryptophan hydroxylase activity and plasma corticosterone levels. Neuroendocrinology 21 (4):338-349; 1976.
3. Azmitia, E.C.; Liao, B.; Chen Y. Increase of tryptophan hydroxylase enzyme protein by dexamethasone in adrenalectomized rat midbrain. J. Neurosci. 13:504-505; 1993.
4. Azmitia, E.C.; McEwen, B.Si Corticosterone regulation of tryptophan hydroxylase in midbrain of the rat. Science 166:1274-1276; 1969.
5. Azmitia, E.C.; McEwen, B.S. Adrenocortical influence on rat brain tryptophan hydroxylase activity. Brain Res. 78:291-302; 1974.
6. Blundell, J.E. Is there a role for serotonin (5-HT) on feeding? Int. J. Obes. 1:15-42; 1977.
7. Blundell, J.E.; Hill, A.J. Nutrition, serotonin, and appetite: Case study in the evolution of a scientific idea. App. 8:183-194; 1987.
8. Bolanos Jim'enez, F.; Manhaes de Castro, R.; Fillion, G. Effect of chronic antidepressant treatment on 5-HT1B presynaptic heteroreceptors inhibiting acetylcholine
release. Neuropharmacology 33:77-81; 1994.
9. Bonvento, G.; Lacombe, P. Serotonin and the cerebral circulation. The Regulation of Cerebral Blood Flow (ed. Phillis, J.W.), Chapter 6:97-112. CRC Press, Boca Raton, FL; 1993.
10. Charney, D.S.; Woods, S.W.; Krystal, J.H.; Heninger, G.R. Serotonin function and human anxiety disorders. Ann. NY Acad. Sci. 600:558-573; 1990.
11. Clifton, P.G.; Barnfield, A.M.C.; Philcox, L. A behavioral profile of fluoxetine- induced anorexia. Psychopharmacology (Berlin) 97:89-95; 1989.
12. Commins, D.L.; Vosmer, G.; Virus, R.M. Biochemical and histological evidence that methylenedioxymethylamphetamine (MDMA) is toxic to neurons in the rat brain. J. Pharmacol Exp. Ther. 241:338-345; 1987.
13. Dourish, C.T. 5-HT receptor subtypes and feeding behavior. Adv. Bioscien. 85:179- 202; 1992.
14.Friedman, P.A.; Kappelman, A.H.; Kaufman, S. Partial purification and characterization of tryptophan hydroxylase from rabbit hindbrain. J. Biol Chem. 247:4165-4173; 1972.
15. Fujita, M.; Shimada, S.; Maeno, H.; Nishimura, T.; Tobyama, M. Cellular localization of serotonin transporter mRNA in the rat brain. Neurosci. Lett. 162:59-62; 1993.
16. Harrington, M.A.; Zhong, P.; Garlow, S.J.; Ciaranello, R.D. Molecular biology of serotonin receptors. J. Clin. Psych. 53: 8-27; 1992.
17. Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P.P.A. International Union of Pharmacology Classification of Receptors for 5-Hydroxytryptamine (Serotonin). Pharmacol. Rev. 46:158-193; 1994.
18. Humphrey, P.P.A. 5-hydroxytryptamine and the pathophysiology of migraine. J. Neurol 238:538-544; 1991.
19. Humphrey, P.P.A.; Hartig, P.; Hoyer, D. A proposed new nomencalture for 5-HT receptors. Trends Pharmacol. Sci. 14:233-236; 1993.
20. Inoue, F.; Hasegawa, H.; Nishimura, M.; Yanagisawa, M.; Ichiyama, A. Distribution of 5-hydroxytryptamine (5-HT) in tissue of a mutant mouse deficient in mast cell. Demonstration of the contribution of mast cells to the 5-HT content in various organs. Agents Actions 16:295-301; 1985.
21. Islam, A.K.; Dougherty, T.; Koch, J.E.; Bodnar, R.J. Naltrexone, serotonin receptor subtype antagonists, and carbohydrate intake in rats. Pharm. Biochem. Behav. 48(1 ): 193-201; 1994.
22. Kennet, G.A. Evidence that mCPP may have behavioral effects mediated by central 5-HT-1C receptors. Br. J. Pharmacol. 94:137-147; 1988.
23. Kennet, G.A.; Curxon, G. Evidence that hypophagia induced mCPP and TFMPP requires 5-HT-1 B receptors and 5-HT-1C receptors: Hypophagia induced by RU24969 only requires 5-HT-1B receptors. Psychopharmacology (Berlin) 96:93-100; 1988.
24. Kennet, G.A.; Dourish, C.T.; Curzon, G. 5-HT-1B agonists produce anorexia at a postsynaptic site. Eur. J. Pharmacol. 141:429-435; 1987.
25. Lee. M.D.; Clifton, P.G. Partial reversal of fluoxetine anorexia by the 5-HT antagonist metergoline. Psychopharmacology (Berlin) 107:359-364; 1992.
26. Liebowitz, S.F.; Weiss, G.F.; Walsh, U.A.; Viswanath, D. Medial hypothalamic serotonin: Role in circadian patterns of feeding and macronutrient selection. Brain Res. 503:132-140; 1989.
27. Luo, S.; Li, E.T.S. Food intake and selection pattern of treated with dexfenfiuramine, fiuoxetine and RU24969. Brain Res. Bull. 24:729-733; 1990.
28. Molliver, M.E. Serotonergic neuronal systems: What their anatomic organization tells us about function. J. Clin. Psychopharmacol. 7:3-23; 1987.
29. Murray, R.K.; Granner, D.K.; Mayes, P.A.; Rodwell, V.W. 1988. Harper's Biochemistry: 491-522. Norwalk, CT: Appleton and Lange.
30. Nobin, A.; Bjorklund, A. Topography of the monoamine neuron system in the human brain as revealed in fetuses. Acta. Physiol. Scand. Suppl. 388: 1-40; 1973.
31. Peroutka, S.J.; Schmidt, A.W.; Sleight, A.J.; Harrington, H.A. Serotonin receptor 'families' in the central nervous system: An overview. Ann. NY Acad. Sci. 600:104-113; 1990.
32. Rattray, M.; Baldessani, S.; Gobbi, M.; Mennini, T.; Samanin, R.; Bendotti, C. P- Chlophenylalanine changes serotonin transporter mRNA levels and expression of the gene product. J. Neurochem. 67 (2):463-472; 1996.
33. Ricaurte, G.; Bryan, G.; Srtraus, L. Hallucinogenic amphetamine selectively destroys brain serotonin nerve terminals. Science 229:986-988; 1985.
34. Ricaurte, G.; Finnegan, K.F.; Nichols, D.E. 3,4-Methylenedioxyethylamphetamine (MDE), a normal analogue of MDMA, produces long-lasting depletion of serotonin in the rat brain. Eur. J. Pharmacol. 137:265-268; 1987.
35. Rubin, R.T.; Poland, R.E.; Lesser, I.M.; Winston, R.A.; BIodgett, N. Neuroendocrine aspects of primary endogenous depression. Arch. Gen. Psychiatry 44:328-336; 1987.
36. Series, H.G.; Molliver, M.E. Immunocytochemical Evidence for Serotonergic Neurotoxicity of N-Ethyl-methylenedioxyamphetamine (MDE). Experimental Neurology 128:50-58; 1994.
37. Schwarz, D.H.; Hernandez, L.; Hoebel, B.G. Tryptophan Increases Extracellular Serotonin in the Lateral Hypothalamus of Food-Deprived Rats. Brain Res. Bull. 25:803- 807; 1990.
38. Shechter, L.E.; Simansky, K.J. 1-(2,5-Dimethoxy-4-iodophenyl)-2-aminopropame (DOI) exerts an anorectic action that is blocked by 5-HT-2 antagonists in rats. Psychopharmacology (Berlin) 94:342-346; 1988.
39. Shor-posner, G.; Grinker, J.A.;Marinescu, C.; Brown, O.; Liebowitz, S.F. Hypothalamic serotonin in the central of meal patterns and macronutrient selection. Brain Res. Bull. 17:663-671; 1986.
40. Siever, L.J. Kahn, R.S.; Lawlor, B.A.; Trestman, R.L. Lawrence, T.L.; Coccaro, E.F. Critical issues in defining the role of serotonin in psychiatric disorders. PharmacoL Rev. 43:509-525; 1991.
41. Suh, J.S.; Weiss, G.F.; Liebowitz, S.F. Impact of central serotonin on macronutrient selection as a function of brain site. Proc. East. Psychol Assoc. 59:55; 1988.
42. West, H.L.; Mark, G.P.; Hoebel, B.G. Effects of conditioned taste aversion on extracellular serotonin in the lateral hypothalamus and hippocampus of freely moving rats. Brain Res. 556:95-100; 1991.
43. Whitaker-Azmitia, P.M.; Peroutka, S.J. The Neuropharmacology of Serotonin. Ann. NY Acad. Sci. 600; 1990.
44. Wilkinson, L.O.; Auerbach, S.B.; Jacobs, B.L. Relationship between dialysate serotonin and raphe unit activity. Soc. Neurosci. Abstr. 14:25; 1988.
45. Young, E.A.; Haskett, R.F.; Murphy-Weinberg, V.; Watson, S.J.; Akil, H. Loss of glucocorticoid fast feedback in depression. Arch. Gen. Psychiatry 48:693-699; 1991. Figures and Table 1 Figure 1. Schematic Representation of the Synthesis of 5-Hydroxytryptamine (Serotonin). The synthesis of serotonin is initiated by the hydroxylation of the amino acid L-tryptophan catalyzed by the enzyme tryptophan hydroxylase and H4 Biopterin, the cofactor. 5-Hydroxtryptophan (5-HTP) is then decarboxylated by L-amino acid decarboxylase enzyme to produce 5-HT (Serotonin) (2). FIGURE 2. Reuptake of Serotonin Via Serotonin (5-HT) Transporter Sites. Serotonin, normally contained in nerve terminals, must be released into the synapse (the space between two neurons) in order to affect other neurons and transduce its effects on appetite, sleep, locomotor activity, and other physiological behaviors. Once the effect has taken place, the extracellular serotonin is taken back into the nerve terminal via serotonin transporter sites. Such sites can be blocked by serotonin reuptake inhibitors, such as paroxetine and Prozac, which compete with serotonin for these transporter sites. Figure 3. Weight of Food-Restricted vs. Ad Libitum Rats. The weights in grams (g) of both the food-restricted and ad libitum (control) rats were recorded daily during the 14 day food-restriction period. The rats were tested in three groups of 8 rats each, and each group is plotted separately in the graph. Mean starting and ending weights for each group are shown in Table 1. Figure 4. Effect of Food-Restriction on Hippocampa/ Serotonin Transporter Site Density. Male Sprague-Dawley rats were maintained on either a food-restricted or an ad libitum (control) diet for 14 days and then sacrificed. Displayed are the results of the three 3H-paroxetine binding assays, a measure of 5-HT transporter sites, for the hippocampus food-restricted and control rats in each of the three experimental groups. In the first and third experimental groups, Hipp 1 and Hipp 3, a significant decrease (p <0.05) of the serotonin transporter site density caused by food-restriction was detected. A significant decrease was not seen in the second experimental group, Hipp 2. Figure 5. Effect of Food-Restriction on Brainstem Serotonin Transporter Site Density. Male Sprague-Dawley rats were maintained on either a food-restricted or an ad libitum (control) diet for 14 days and then sacrificed. Displayed are the results of 3H- paroxetine binding assays, a measure of the 5-HT transporter site density, for the brainstem from food-restricted and control rats in each of the three experimental groups. In the first and third experimental groups, Brainstem 1 and Brainstem 3, a significant decrease (p <0.05) of the serotonin transporter site density caused by food-restriction was detected. A significant decrease was not seen in the second experimental groups. Figure 6. Effect of Food-Restriction on Cortica/ Serotonin Transporter Site Density. Male Sprague-Dawley rats were maintained on either a food-restricted or an ad libitum (control) diet for 14 days and then sacrificed. Results from food-restricted subjects were compared to ad libitum subjects. Food-restriction did not result in a significant decrease (p <0.05) in serotonin transporter sites in any of the three experiments. This figure displays the amount of 3H-paroxetine bound to the cortex membrane homogenates in both the control and food-restricted subjects. The greater the amount of binding, the greater the serotonin transporter site density. Figure 7. Effect of Food-Restriction on Striatal Serotonin Transporter Site Density. Male Sprague-Dawley rats were maintained on either a food-restricted or an ad libitum (control) diet for 14 days and then sacrificed. Results from food-restricted subjects were compared to ad libitum subjects. Food-restriction produced a significant decrease (p- <0.05) in serotonin transporter sites only in the third experimental group, Striaturn 3. No significant decrease was detected in the first two experimental groups, Striaturn 1 and Striaturn 2. Although the percentage difference between the subjects seemed large, especially in group 1, the large standard deviation in the ad libitum values compromised the significance.
Figure 8. Schematic Representation of the Hypothalamic-Pituitary-Adrenal Axis. The release of corticosterone is regulated via a negative feedback loop where the greater amount of plasma corticosterone present, the less is produced due to an inhibitory pathway created by the increased level. Corticosterone-releasing hormone (CRH), which is released by the hypothalamus, causes the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH, in turn, increases the amount of cholesterol that is converted to pregnenolone in the adrenal cortex. This then leads to the heightened production of corticosterone. Serotonin has a putative neurological role in this system through its influence on the hypothalamus and the release of CRH (29). COMPARISON OF THE STARTING AND ENDING MEAN BODY WEIGHTS OF THE SUBJECT GROUPS TABLE 1. Weight Change of Subjects During Food-Restriction. The weights of each of the 24 rats were recorded prior to and during the period of food-restriction, as well as on the day of sacrifice. Averages of beginning weights (prior to food-restriction) and ending weights (on the day of sacrifice) of food-restricted and control groups wee calculated and the average ending weights of each group are displayed as a percentage of the average beginning weights of the respective group.
Copyright to this work is retained by the author[s]. Permission is granted for the noncommercial reproduction of the complete work for educational or research purposes.
Collecting Data with the Blood Pressure Sensor This sensor can be used with the following interfaces to collect data: Blood Pressure • Vernier LabQuestTM as a standalone device or with a computer (Order Code BPS-BTA) Here is the general procedure to follow when using the Blood Pressure Sensor: The Vernier Blood Pressure Sensor is used to 1. Connect the Blood Pressure Sensor to th
January 1, 2014 Complimentary Consultation In our continuing quest to introduce more people (including children) to the benefits of chiropractic care we are offering a complimentary consultation to see if chiropractic can help. Call the office for an appointment and bring in this page in order to receive your FREE consultation. Call now: 856-778-8688 Enjoy the benefi
 Chronic Food-Restriction Decreases Brain
Chronic Food-Restriction Decreases Brain  PVN- Paraventricular nucleus of the hypothalamus
PVN- Paraventricular nucleus of the hypothalamus 10g of food per day and the remaining 4 rats were fed ad libitum (intake is approximately 20-30g of food per day) for 14 days. Therefore, 12 rats in total were food-restricted and 12 were fed ad libitum. At the end of the 14 days, all of the subjects were sacrificed. The hippocampus, striaturn, brainstem, and cortex regions of the brain were removed, frozen, and stored. The density of 5-HT transporter sites (the sites at which released 5-HT is taken back into the cell) was estimated by measuring paroxetine-binding by membranes from cells in these brain regions. Paroxetine is known to bind to 5-HT transporter sites.
10g of food per day and the remaining 4 rats were fed ad libitum (intake is approximately 20-30g of food per day) for 14 days. Therefore, 12 rats in total were food-restricted and 12 were fed ad libitum. At the end of the 14 days, all of the subjects were sacrificed. The hippocampus, striaturn, brainstem, and cortex regions of the brain were removed, frozen, and stored. The density of 5-HT transporter sites (the sites at which released 5-HT is taken back into the cell) was estimated by measuring paroxetine-binding by membranes from cells in these brain regions. Paroxetine is known to bind to 5-HT transporter sites. decreased levels of 5-HT transporter sites in the hippocampus, brainstem, and striatum. Such information could be useful in an animal model in the study of bulimia, obesity, and depression, disorders which are effectively treated with 5-HT reuptake inhibitors.
decreased levels of 5-HT transporter sites in the hippocampus, brainstem, and striatum. Such information could be useful in an animal model in the study of bulimia, obesity, and depression, disorders which are effectively treated with 5-HT reuptake inhibitors. to vomit. Instead, when an unfavorable substance has been ingested, a conditioned taste aversion ensues where the rodent will no longer ingest that substance. A conditioned taste aversion to saccharin solution can be instilled in rodents. When a saccharin solution, to which a conditioned taste aversion had been instilled, was orally infused into rodents, elevated levels of 5-HT were observed in the LH. The same elevated levels were also noted in rodents receiving injections of lithium chloride (LiCl), which is illness inducing. (42).
to vomit. Instead, when an unfavorable substance has been ingested, a conditioned taste aversion ensues where the rodent will no longer ingest that substance. A conditioned taste aversion to saccharin solution can be instilled in rodents. When a saccharin solution, to which a conditioned taste aversion had been instilled, was orally infused into rodents, elevated levels of 5-HT were observed in the LH. The same elevated levels were also noted in rodents receiving injections of lithium chloride (LiCl), which is illness inducing. (42). binding to obtain the specific binding value of 3H-paroxetine. These tubes were then vortexed and incubated for 15 minutes at room temperature. Subsequently, 200 ul of 3H-paroxetine buffer was added to each tube. The samples were then incubated for 120 minutes at room temperature. Following this final incubation, the samples were filtered over 0.1% polyethyleneimine (PEI) coated filters utilizing a Brandell cell harvester. PEI prevents any free ligand from binding to the filter. Each glass fiber filter was then placed into a glass test tube which was filled with 5 ml of Ecoscint A scintillation fluid. The tubes were then placed into a scintillation counter to count the amount of 3H-paroxetine bound to the rat brain membrane homogenates.
binding to obtain the specific binding value of 3H-paroxetine. These tubes were then vortexed and incubated for 15 minutes at room temperature. Subsequently, 200 ul of 3H-paroxetine buffer was added to each tube. The samples were then incubated for 120 minutes at room temperature. Following this final incubation, the samples were filtered over 0.1% polyethyleneimine (PEI) coated filters utilizing a Brandell cell harvester. PEI prevents any free ligand from binding to the filter. Each glass fiber filter was then placed into a glass test tube which was filled with 5 ml of Ecoscint A scintillation fluid. The tubes were then placed into a scintillation counter to count the amount of 3H-paroxetine bound to the rat brain membrane homogenates. displayed a decrease in the 5-HT transporter sites in food-restricted subjects. The difference was 21% between food-restricted and control subjects, and specifically there was 262 fmol/mg average specific 3H-paroxetine binding in the control subjects compared to 207 fmol/mg in the food-restricted subjects. The p value for the third experimental group was 0.01.
displayed a decrease in the 5-HT transporter sites in food-restricted subjects. The difference was 21% between food-restricted and control subjects, and specifically there was 262 fmol/mg average specific 3H-paroxetine binding in the control subjects compared to 207 fmol/mg in the food-restricted subjects. The p value for the third experimental group was 0.01.
 (CRH) from the hypothalamus to induce the basophilic cells of the anterior pituitary to produce, as well as release, adrenocorticotropic hormone (ACTH). Several neurotransmitters, one of which is serotonin, indirectly stimulate the release of CRH at this point of the cycle via 5-HT1A and 5-HT2A/2C receptors (29). Glucocorticoids affect the
serotonin system by stimulating 5-HT synthesis and stress-induced tryptophan hydroxylase activity (3,4,5). Further correlation is observed through the fact that glucocorticoids are also affected by food-restriction. Plasma corticosterone (the major glucocorticoid in rats) levels have been shown to be elevated preceding the scheduled daily meal in food-restricted rats (1). Since abnormalities in the HPA axis have been shown to be correlated to depression (35,45) and one of the major aberrant behavioral effects of disturbances in the neurotransmission of 5-HT is depression (10,40), experimentation regarding the correlation between the effects of food-restriction on both systems could prove useful.
(CRH) from the hypothalamus to induce the basophilic cells of the anterior pituitary to produce, as well as release, adrenocorticotropic hormone (ACTH). Several neurotransmitters, one of which is serotonin, indirectly stimulate the release of CRH at this point of the cycle via 5-HT1A and 5-HT2A/2C receptors (29). Glucocorticoids affect the
serotonin system by stimulating 5-HT synthesis and stress-induced tryptophan hydroxylase activity (3,4,5). Further correlation is observed through the fact that glucocorticoids are also affected by food-restriction. Plasma corticosterone (the major glucocorticoid in rats) levels have been shown to be elevated preceding the scheduled daily meal in food-restricted rats (1). Since abnormalities in the HPA axis have been shown to be correlated to depression (35,45) and one of the major aberrant behavioral effects of disturbances in the neurotransmission of 5-HT is depression (10,40), experimentation regarding the correlation between the effects of food-restriction on both systems could prove useful.


 Figures and Table 1
Figures and Table 1
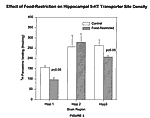

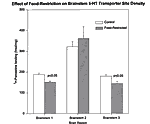
 Figure 3. Weight of Food-Restricted vs. Ad Libitum Rats. The weights in grams (g) of
Figure 3. Weight of Food-Restricted vs. Ad Libitum Rats. The weights in grams (g) of 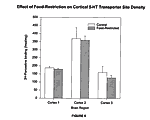

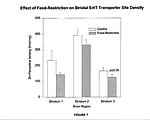

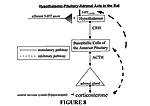 Figure 6. Effect of Food-Restriction on Cortica/ Serotonin Transporter Site Density.
Figure 6. Effect of Food-Restriction on Cortica/ Serotonin Transporter Site Density. 

 Figure 8. Schematic Representation of the Hypothalamic-Pituitary-Adrenal Axis. The release of corticosterone is regulated via a negative feedback loop where the greater amount of plasma corticosterone present, the less is produced due to an inhibitory pathway created by the increased level. Corticosterone-releasing hormone (CRH), which is released by the hypothalamus, causes the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH, in turn, increases the amount of cholesterol that is converted to pregnenolone in the adrenal cortex. This then leads to the heightened production of corticosterone. Serotonin has a putative neurological role in this system through its influence on the hypothalamus and the release of CRH (29).
Figure 8. Schematic Representation of the Hypothalamic-Pituitary-Adrenal Axis. The release of corticosterone is regulated via a negative feedback loop where the greater amount of plasma corticosterone present, the less is produced due to an inhibitory pathway created by the increased level. Corticosterone-releasing hormone (CRH), which is released by the hypothalamus, causes the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH, in turn, increases the amount of cholesterol that is converted to pregnenolone in the adrenal cortex. This then leads to the heightened production of corticosterone. Serotonin has a putative neurological role in this system through its influence on the hypothalamus and the release of CRH (29).