La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Alford.bios.uic.edu
The Journal of Neuroscience, May 1, 1998, 18(9):3138–3146
G-Protein-Coupled Modulation of Presynaptic Calcium Currents and Transmitter Release by a GABA Receptor Tomoyuki Takahashi, Yoshinao Kajikawa, and Tetsuhiro Tsujimoto Department of Neurophysiology, University of Tokyo Faculty of Medicine, Tokyo 113, Japan
Presynaptic GABA receptors play a regulatory role in central
(GDPS) abolished the effect of baclofen on both presynaptic
synaptic transmission. To elucidate their underlying mechanism
calcium currents and EPSCs. The nonhydrolyzable GTP analog
of action, we have made whole-cell recordings of calcium and
guanosine 5Ј-O-(3-thiotriphosphate) (GTP␥S) suppressed pre-
potassium currents from a giant presynaptic terminal, the calyx
synaptic calcium currents and occluded the effect of baclofen
of Held, and EPSCs from its postsynaptic target in the medial
on presynaptic calcium currents and EPSCs. Photoactivation of
nucleus of the trapezoid body of rat brainstem slices. The
GTP␥S induced an inward rectifying potassium current at the
GABA receptor agonist baclofen suppressed EPSCs and pre-
calyx of Held, whereas baclofen had no such effect. We con-
synaptic calcium currents but had no effect on voltage-
clude that presynaptic GABA receptors suppress transmitter
dependent potassium currents. The calcium current–EPSC re-
lationship measured during baclofen application was similar to
that observed on reducing [Ca 2ϩ] , suggesting that the pre-
synaptic inhibition generated by baclofen is caused largely by
receptor; presynaptic inhibition; G-
the suppression of presynaptic calcium influx. Presynaptic
protein; calcium currents; inwardly rectifying potassium cur-
loading of the GDP analog guanosine-5Ј-O-(2-thiodiphosphate)
rents; the calyx of Held; presynaptic recording
GABAB receptors are widely distributed in the presynaptic and
GABAB receptors as well as metabotropic glutamate receptors
postsynaptic membranes of vertebrate central neurons, and they
and adenosine receptors (Barnes-Davies and Forsythe, 1995).
modulate synaptic transmission by either suppressing transmitter
Here we demonstrate that the G-protein-coupled inhibition of
release or hyperpolarizing postsynaptic cells (Thompson et al.,
calcium channels underlies the GABAB receptor-mediated pre-
1993; Kaupmann et al., 1997). At neuronal somata, GABAB
receptors are known to activate G-proteins, thereby enhancing
inwardly rectifying potassium channels (Andrade et al., 1986;
MATERIALS AND METHODS
Sodickson and Bean, 1996) or suppressing calcium channels
Preparation and solutions. Transverse slices of the superior olivary com-
(Dolphin and Scott, 1987; Scholtz and Miller, 1991; Mintz and
plex were prepared from 14- to 19-d-old Wistar rats killed by decapitation
Bean, 1993). Compared with the wealth of information on the
under halothane anesthesia. The medial nucleus of trapezoid body
(MNTB) neurons and calyces were viewed with a 40ϫ, 63ϫ (Zeiss), or
60ϫ (Olympus Optical, Tokyo, Japan) water immersion lens attached to
known about their presynaptic mechanism of action. In particu-
an upright microscope (Axioskop, Zeiss). Each slice was perfused with
lar, it is not known whether the effector of presynaptic GABAB
artificial CSF (aCSF) containing 120 mM NaCl, 2.5 mM KCl, 26 mM
receptors is a potassium channel (Saint et al., 1990; Thompson
NaHCO3, 1.25 mM NaH2PO4, 2 mM CaCl2, 1 mM MgCl2, 10 mM
and Gahwiler, 1992), a calcium channel (Scholtz and Miller, 1991;
glucose, 0.5 mM myo-inositol, 2 mM sodium pyruvate, 0.5 mM ascorbic
Pfrieger et al., 1994; Wu and Saggau, 1995; Dittman and Regehr,
M lactic acid, pH 7.4, with 5% C O2 and 95% O2. To isolate
Ca 2ϩ currents, 10 mM tetraethylammonium (TEA) chloride and 1 M
1996, 1997), or exocytotic machinery downstream of calcium
tetrodotoxin (TTX) were included in the aCSF. The postsynaptic patch
influx (Scanziani et al., 1992; Dittman and Regehr, 1996;
pipette was filled with a solution (A) containing 97.5 mM potassium
Rohrbacher et al., 1997). Also, an involvement of G-proteins in
gluconate, 32.5 mM KCl, 10 mM HEPES, 5 mM EGTA, and 1 mM MgCl2,
pH adjusted to 7.4 with KOH. N-(2,6-diethylphenylcarbamoylmethyl)-
B receptor-mediated presynaptic inhibition remains to be
triethyl-ammonium bromide (QX314, 5 mM) was included in the postsyn-
directly demonstrated (Thompson et al., 1993). The calyx of Held
aptic pipette solution to suppress action potential generation when aCSF
is an ideal preparation for directly testing these issues using
did not contain TTX. For recording EPSCs, the aCSF routinely con-
patch-clamp techniques (Forsythe, 1994; Borst et al., 1995;
tained bicuculline (10 M) and strychnine (0.5 M) to block spontaneous
Takahashi et al., 1996). The presynaptic terminal possesses
inhibitory synaptic currents. For recording presynaptic calcium currents
(IpCa), the presynaptic pipette was filled with a solution (B) containing
110 mM CsCl, 40 mM HEPES, 0.5 mM EGTA, 1 mM MgCl2, 2 mM ATP,
0.5 mM GTP, 12 mM Na2 phosphocreatinine, and 10 mM TEA, pH
Received Nov. 26, 1997; revised Feb. 11, 1998; accepted Feb. 12, 1998.
adjusted to 7.4 with CsOH. Presynaptic potassium currents were re-
This work was supported by the “Research for the Future” Program by The Japan
corded with solution A. The presynaptic pipette solutions routinely
Society for the Promotion of Sciences. We thank M. Farrant, I. D. Forsythe, T.
contained 2 mM ATP (ATP-Mg salt), 12 mM phosphocreatinine, and 0.5
Manabe, and K. Kobayashi for critically reading this manuscript. We are also
mM GTP, unless noted otherwise. For paired recordings 10 mM potas-
grateful to R. Y. Tsien and V. Lev-Ram for their technical advice on the caged
compound photolysis system and to Novartis Pharma (Basel, Switzerland) for the
sium glutamate or cesium glutamate (equimolar replacement of KCl or
CsCl) was also included in the presynaptic pipette solution (Borst et al.,
Correspondence should be addressed to Tomoyuki Takahashi, Department of
Neurophysiology, University of Tokyo Faculty of Medicine, Tokyo 113, Japan. Recording and data analysis. Whole-cell patch-clamp recordings were
Copyright 1998 Society for Neuroscience 0270-6474/98/183138-09$05.00/0
made from MNTB neurons, presynaptic calyces, or simultaneously from
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
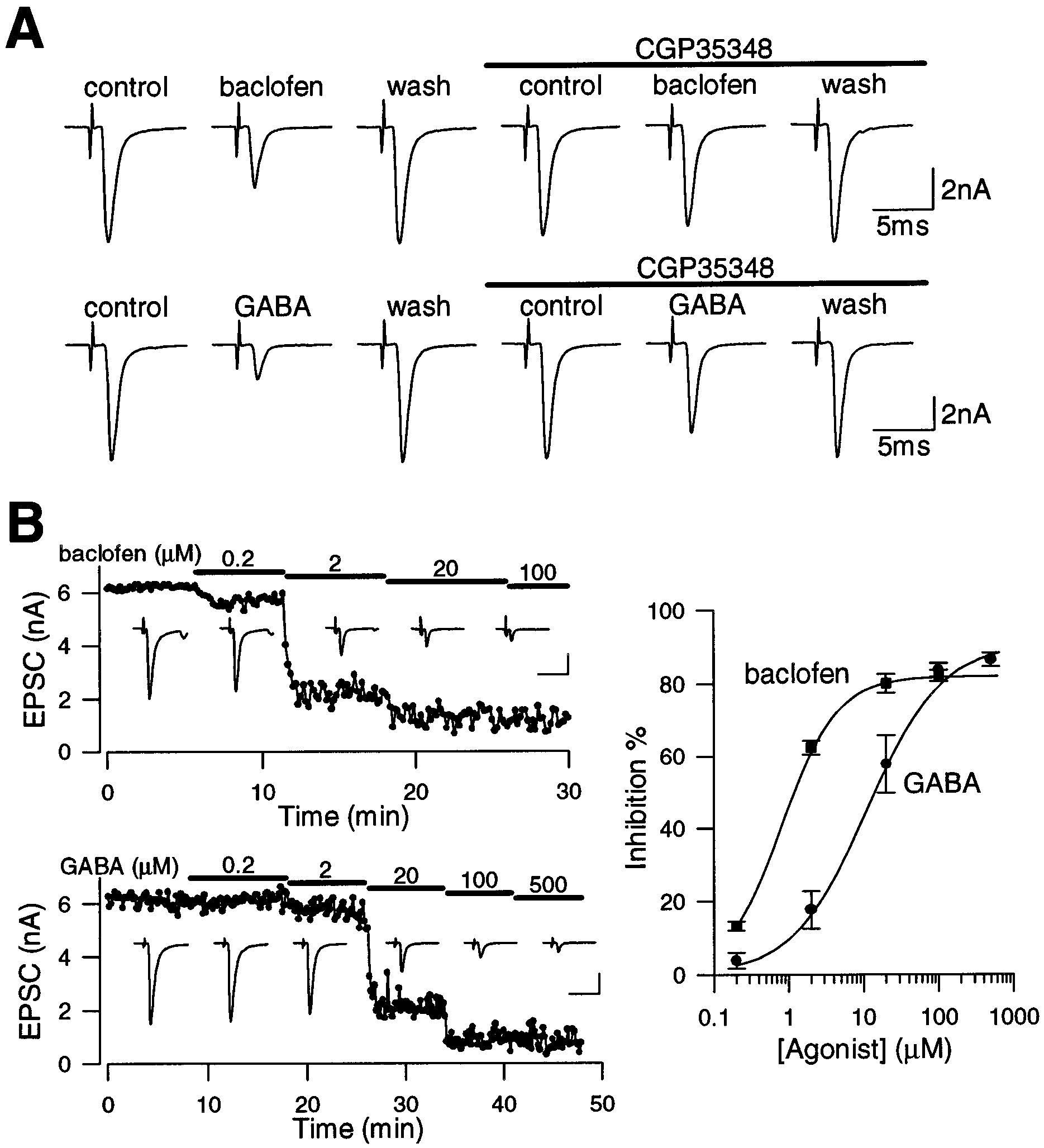
J. Neurosci., May 1, 1998, 18(9):3138–3146 3139 Figure 1. Inhibitory effects of GABAB receptor agonists on EPSCs. EPSCs were evoked by extracellular stimulation. A (top row), Reversible inhibition
of EPSCs by baclofen (2 M) and attenuation of the baclofen effect by CGP35348 (100 M) in an MNTB neuron. Bottom row, Inhibitory effect of GABA
(20 M) and its attenuation by CGP35348 in another MNTB neuron. The magnitude of inhibition by CGP35348 on the effect of baclofen and GABA
was 82.8 Ϯ 2.3% (n ϭ 3) and 60.7 Ϯ 11% (n ϭ 3), respectively. B, Dose-dependent suppression of EPSCs by baclofen and GABA. Cumulative
dose-dependent effects of baclofen (top) and GABA (bottom) on the amplitude of EPSCs recorded from MNTB neurons. Sample records from
individual MNTB neurons are shown in the inset. Calibration: 2 nA, 10 msec. The curves fitted to data points derived from the following equation:
magnitude of inhibition (%) ϭ maximal inhibition (%)/[1 ϩ (IC50/agonist concentration)n]. For baclofen and GABA, maximal inhibition was 82.0 and
90.8%, IC50 was 0.77 and 9.97 M, and Hill coefficient (n) was 0.90 and 1.21, respectively. Magnitude of EPSC suppression by 20 M baclofen was 80.1 Ϯ
2.6% at the cumulative-dose application (n ϭ 4), which was not significantly different ( p ϭ 0.13; Student’s t test) from that at the single-dose application
both structures (Takahashi et al., 1996). EPSCs were evoked at 0.1 Hz by
recording was typically 10 –20 M⍀ and was compensated by 60–90%.
extracellular stimulation of presynaptic axons near the midline of a slice
Current or potential recordings were made with a patch-clamp amplifier
with a bipolar platinum electrode (Barnes-Davies and Forsythe, 1995) in
(Axopatch 200B, Axon Instruments, Foster City, CA). Unless noted
a relatively thick slice (250 m) or by presynaptic action potentials or
otherwise, records were low-pass-filtered at 2.5–20 kHz and digitized at
Ca 2ϩ currents elicited by a whole-cell pipette in thin slice (150 m). The
5–50 kHz by a CED 1401 interface (Cambridge Electronic Design). Leak
electrode resistances were 4 –7 M⍀ for the postsynaptic pipette and 6–10
currents were subtracted for presynaptic currents by a scaled pulse
M⍀ for the presynaptic pipette. The series resistance of presynaptic
divided by n (P/N) protocol. The liquid junctional potential between the
3140 J. Neurosci., May 1, 1998, 18(9):3138–3146
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
Figure 2. Baclofen-induced suppression of EPSCs is
blocked by GDPS. Simultaneous presynaptic and
postsynaptic recordings at the calyx–MNTB synapse.
EPSCs were evoked by action potentials elicited by a
depolarizing current pulse (2–10 msec) applied to a
calyx through a whole-cell patch pipette. The postsyn-
aptic holding potential was Ϫ70 mV. A, Reversible
suppression of EPSCs by baclofen (20 M). B, Block-
ing effect of GDPS (tri-lithium salt, 3 mM) in the
pipette on baclofen-induced suppression of EPSCs (a,b). A lower concentration of GDPS (0.2 mM) did not
prevent the effect of baclofen (data not shown). After
the pipettes were retracted, a second paired recording
was made from the same structures with a presynaptic
pipette containing GTP (0.5 mM) instead of GDPS.
Baclofen clearly suppressed the EPSCs (c, d), which
gradually recovered after washout (e). Complete re-
covery of EPSCs took 5–10 min (Fig. 7). When LiCl (9
mM) was included in the presynaptic pipette the ba-
clofen effect was not attenuated (not shown). The
amplitudes of EPSCs were normalized against the
mean of the first seven (with GDPS) or six (with
GTP) data points before baclofen application in each
experiment; the data point represents means and the
error bars represent SEMs derived from paired re-
cording experiments at three different synapses. Ver-
tical calibration scales indicate 80 mV for presynaptic
membrane potentials (A and B) and 1.25 nA (A) or 0.6
nA ( B) for EPSCs. Scale bars, 10 msec.
pipette solution and aCSF was ϩ7.5 mV for solution A and ϩ3.3 mV for
reported previously (Barnes-Davies and Forsythe, 1995), bath-
solution B. The value of reversal potentials (see Fig. 6C) was corrected
EPSCs in a reversible manner (Fig. 1A). This baclofen effect was
switching superfusates by solenoid valves. Caged GTP␥S [S-(DMNPE-
detectable at 0.2 M, increased dose-dependently, and reached a
caged) GTP␥S; Molecular Probes, Eugene, OR] was applied at 38 M
maximal at ϳ20 M (Fig. 1B). Similarly, the inhibitory transmit-
into calyces by dialysis from whole-cell pipettes. Care was taken to
ter GABA suppressed EPSCs (Fig. 1). The 50% inhibitory con-
protect the compound from short wavelength light during this procedure.
A flash of light was given from a mercury lamp light source (50 W)
50 ) of baclofen was estimated from the dose –
through a filter (360 Ϯ 20 nm) by opening a shutter for a given period
response curve to be 0.8 M, whereas that for GABA was 10 M
(2– 4 sec). Application of the light flash without loading caged compound
(Fig. 1B). Thus baclofen was about 10 times more potent than
had no effect on the synaptic transmission or IpCa under normal exper-
GABA in inhibiting EPSCs at this synapse. The inhibitory effects
imental conditions, although an excessive illumination sometimes in-
of both baclofen and GABA were largely attenuated by the
duced a transient potentiation of IpCa or an increase in the frequency of
spontaneous synaptic currents. Experiments were carried at room tem-
GABAB receptor antagonist CGP35348 (100 M) (Fig. 1A), in-
dicating that the effects of baclofen and GABA were indeed
Presynaptic inhibition mediated by GABA receptors at Involvement of G-proteins in GABA receptor- the calyx of Held mediated presynaptic inhibition
A single extracellular stimulation evoked a large and rapidly
An EPSC was evoked by a presynaptic action potential in a
decaying EPSC in a principal cell of the medial nucleus of
simultaneous whole-cell recording from the calyx of Held and a
trapezoid body (MNTB) under whole-cell voltage clamp. As
target MNTB cell (Fig. 2A). Baclofen suppressed the EPSC
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3141
are involved in the GABAB receptor-mediated presynaptic
Inhibition of presynaptic calcium currents by baclofen To identify an effector of the presynaptic GABAB receptor, we
first examined whether presynaptic calcium currents (IpCa) could
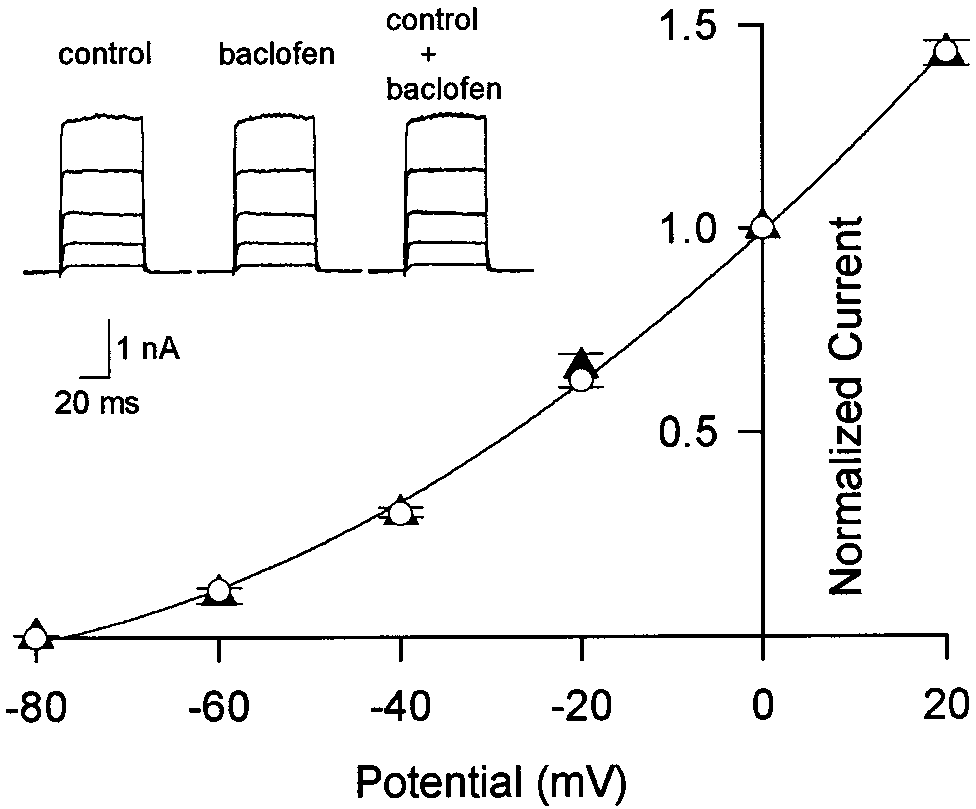
be modulated by baclofen. As illustrated in Figure 3A, baclofen
slowed activation kinetics of IpCa and reduced its amplitude.
When measured at the peak of the control current (1.3 msec from
onset) at Ϫ10 mV, the magnitude of IpCa suppression was 38.0 Ϯ
3.8% (n ϭ 6). The baclofen-induced suppression of IpCa was not
associated with a shift in the current–voltage (I–V) relationship
(Fig. 3C). As shown in Figure 3A,B, after a 10 msec depolarizing
pulse (to Ϫ10 mV) IpCa deactivated exponentially with a fast
time constant (0.14 Ϯ 0.03 msec; n ϭ 8). Baclofen had no effect on
this deactivation time constant (0.14 Ϯ 0.05 msec after baclofen).
This suggests that baclofen has little effect on the presynaptic
Ca2ϩ channel open time. These characteristics of the baclofen-
induced inhibition of IpCa are similar to those reported for
somatic Ca2ϩ currents (Dolphin and Scott, 1987; Scholtz and
Miller, 1991; Mintz and Bean, 1993; Lambert and Wilson, 1996).
To study further the involvement of G-proteins in the baclofen-
induced suppression of IpCa, caged GTP␥S (38 M) was loaded
into a calyx through a whole-cell patch pipette (Fig. 4A). After it
was confirmed that baclofen reversibly suppressed IpCa (a–c), a
flash of ultraviolet light (UV, 340–380 nm) was applied for 2–4
sec (arrow) to induce a photo-release of the caged GTP␥S com-
pound. After the flash, IpCa gradually diminished in amplitude
Figure 3. Suppression of presynaptic Ca 2ϩ currents by baclofen. The
and slowed in its rising phase (c, d). After IpCa amplitude
calyx was voltage-clamped at Ϫ80 mV, and IpCa was evoked by a 10 msec
reached a steady level, a second application of baclofen no longer
depolarizing pulse. In this experiment, [Ca 2ϩ]o was reduced to 1 mM
attenuated IpCa (d, e). In agreement with this result, when
to allow better voltage-clamp performance. A, IpCa induced in a calyx by
a depolarizing voltage step to Ϫ10 mV in the absence and presence of
␥S (200 M) was included in the presynaptic whole-cell
baclofen (20 M, superimposed). B, The tail currents are normalized at
pipette, IpCa exhibited a similarly slow rise, and baclofen had no
the peak and superimposed. C, Current–voltage relationships of IpCa
significant effect on the current amplitude (99.8 Ϯ 1.6%; n ϭ 4)
before (open circles) and after ( filled circles) baclofen application. Mean
(Fig. 4B). When GDPS (3 mM) was included in the pipette, IpCa
values Ϯ SEMs obtained from six calyces are shown.
had a normal rise time, but baclofen was again ineffective on
IpCa (96.9 Ϯ 1.2%; n ϭ 5) (Fig. 4C). These results indicate that
without affecting the presynaptic action potential. The magnitude
the inhibitory effect of baclofen on the presynaptic calcium cur-
of suppression of EPSCs by baclofen (20 M) was 78.5 Ϯ 0.71%
(mean Ϯ SEM; n ϭ 4 cells), which was comparable with that for
the extracellularly evoked EPSCs (Fig. 1 and legend). The pre-
Lack of baclofen effect on presynaptic
synaptic action potential had a peak amplitude of 90.5 Ϯ 12 mV
potassium currents
and a half-width of 0.76 Ϯ 0.19 msec (n ϭ 4 calyces), which
We next examined whether baclofen might modulate potassium
remained at 99.0 Ϯ 2.4% and 113 Ϯ 25%, respectively, during
currents. Voltage-dependent outward potassium currents were
baclofen application. Baclofen had no effect on the presynaptic
evoked by depolarizing a presynaptic terminal in the presence of
membrane potential or conductance (see below).
TTX (1 M) (Forsythe, 1994). As illustrated in Figure 5, the
To directly address an involvement of presynaptic G-proteins
potassium current before and after baclofen application was
in the action of baclofen, the GDP analog guanosine-5Ј-O-(2-
nearly identical at all voltages examined. Thus, GABAB receptors
thiodiphosphate) (GDPS, 3 mM) was included in the presynaptic
do not seem to be coupled with voltage-gated potassium channels
whole-cell pipette. In this condition, baclofen no longer sup-
pressed EPSCs (102 Ϯ 2.6%; n ϭ 4 pairs) (Fig. 2B). After the
In neuronal somata, baclofen enhances inwardly rectifying
whole-cell pipette containing GDPS was retracted, another
potassium currents by activating G-proteins (Andrade et al., 1986;
paired recording was made again at the same synapse, this time
Sodickson and Bean, 1996). We examined whether baclofen
with a presynaptic pipette solution containing GTP. Baclofen
might similarly enhance the inward rectifying potassium current
clearly suppressed EPSCs by 67 Ϯ 14% (n ϭ 3 pairs after GDPS
at the presynaptic terminal. As illustrated in Figure 6A, baclofen
washout) (Fig. 2B). Thus presynaptic GDPS blocked the effect
applied at Ϫ70 mV holding potential had no effect on the holding
of baclofen in a reversible manner. Similarly, when the nonhydro-
current or the membrane conductance (98.5 Ϯ 1.8%; n ϭ 9)
lyzable GTP analog guanosine 5Ј-O-(3-thiotriphosphate)
measured by a ramp command voltage pulse (Fig. 6C). The
(GTP␥S, 200 M) was included in the presynaptic whole-cell
inwardly rectifying potassium current is known to be blocked by
pipette, baclofen had no effect on EPSCs (105 Ϯ 7.6%; n ϭ 3
a low concentration of Ba2ϩ (Hagiwara et al., 1978). Bath-
pairs; data not shown). These results indicate that the effect of
application of Ba2ϩ (100 M) caused a small inward current
baclofen on EPSCs is indeed presynaptic and that G-proteins
accompanied by a slight decrease in membrane conductance (to
3142 J. Neurosci., May 1, 1998, 18(9):3138–3146
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
Figure 4. Block of baclofen-induced IpCa sup-
pression by GTP␥S or GDPS. A, Occlusion of
baclofen effect by GTP␥S. IpCa was evoked in a
calyx by a 20 msec depolarizing step from Ϫ70
mV to Ϫ13 mV. Baclofen (20 M) suppressed
IpCa, which recovered partially (a–c, superim-
posed). After a light flash given at an arrow for 2
sec, IpCa diminished gradually (c, d). A second
application of baclofen after the flash had no
effect on IpCa (d, e). Essentially the same result
was obtained in two other calyces. B, C, Little
effect of baclofen on IpCa (evoked by a 20 msec
depolarizing pulse from Ϫ80 mV to Ϫ10 mV)
was observed in the presence of GTP␥S (200 M,
B) or GDPS (3 mM, C) in the presynaptic pi-
pette. B and C are from different calyces. A
similar result was obtained in another calyx for
84.5 Ϯ 4.6%; n ϭ 6) (Fig. 6A), suggesting that the inwardly
rectifying channels might weakly contribute to the resting con-ductance of the presynaptic terminal. After a calyx was loadedwith caged GTP␥S, photo-release of GTP␥S by a flash (Fig. 6B,
arrow) induced a prominent outward current accompanied by anincrease in membrane conductance. After the outward currentreached a steady level, subsequent application of Ba2ϩ (100 M)
largely abolished this current. When Ba2ϩ was washed out, theoutward current gradually recovered, with an increase in mem-brane conductance (not shown). The Ba2ϩ-sensitive current in-duced by GTP␥S was extracted as a difference current before and
after the Ba2ϩ application (Fig. 6B, a and b). This currentrectified inwardly and reversed at Ϫ92 Ϯ 1.1 mV (n ϭ 4) close to
the theoretical potassium equilibrium potential (99.5 mV; Fig. 6C,arrow), indicating that it is a G-protein-activated inwardly recti-fying potassium current (GIRK) (Kubo et al., 1993). Thus GIRKis present in the presynaptic terminal but cannot be activated byGABAB receptors. Figure 5. Lack of baclofen effect on voltage-gated potassium currents.
Similar to Ba2ϩ, extracellular Csϩ blocks inwardly rectifying
Inset, Outward potassium currents evoked by 20 mV depolarizing steps
potassium currents (Hagiwara et al., 1976; Sodickson and Bean,
from the holding potential of Ϫ80 mV to ϩ20 mV in the presence of TTX
before (left) and after (middle) baclofen (20 M) application. The super-
1996) as well as the inwardly rectifying cationic currents Ih
imposed traces before and after baclofen application overlapped almost
(Halliwell and Adams, 1982; Takahashi, 1990). Bath-application
completely (right). The amplitude of the potassium current was normal-
of Ba2ϩ or Csϩ (both at 1 mM) had no effect on EPSCs evoked
ized against the value at 0 mV and mean Ϯ SEMs of five calyces before
extracellularly (Fig. 7). Baclofen applied in the presence of Ba2ϩ
(open circles) and after ( filled triangles) baclofen application are plotted
or Csϩ suppressed EPSCs to a similar extent as in control: 72.5 Ϯ
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3143 Figure 6. Effects of baclofen and GTP␥S on presynaptic
holding current and membrane conductance. A calyx was
voltage-clamped at the holding potential of Ϫ70 mV, and a
ramp command voltage from Ϫ50 to Ϫ130 mV (C, top left)
was applied every 20 sec. A, Baclofen (20 M) had no effect
on the holding current or input conductance. Ba 2ϩ (100
M) caused a slight inward current associated with a de-
crease in conductance in the same calyx. B, In another
calyx, photo-release of GTP␥S by a UV flash (arrow)
induced an outward current accompanied by an increase in
input conductance. This current was suppressed by Ba 2ϩ
(100 M, b). Application of the light flash without loading
caged compound had no effect on the holding current or
membrane conductance. The outward current was not ob-
served after GTP␥S photolysis with the Csϩ-based internal
solution for IpCa recordings (Fig. 4A). C, Currents (a, b,bottom) corresponding to a command voltage (top) after
photolysis of caged GTP␥S compound before (a) and after
( b) application of Ba 2ϩ. Right, Ba 2ϩ-sensitive current ex-
tracted as a difference current (a–b). Arrow indicates the-
oretical equilibrium potential for potassium ions calculated
from the internal and external potassium activities. The
difference current between before and after photolysis had
a similar reversal potential, but inward rectification was less
prominent (data not shown). Membrane potential was cor-
rected for the liquid junction potential between the exter-
nal and internal solution (ϩ7.5 mV) for this current–
voltage relationship. The data in this figure were low-pass-
filtered at 100 Hz and sampled at 1 kHz.
4.3% in control (n ϭ 8), 72.6 Ϯ 2.0% in Ba2ϩ (n ϭ 4), and 74.8 Ϯ
The result was essentially the same when the charge instead of the
1.5% in Csϩ (n ϭ 4), respectively. These results suggest further
peak amplitude for IpCa and EPSCs was compared. Thus, the
that neither GIRK nor Ih is involved in the GABAB receptor-
baclofen-induced suppression of EPSCs can be explained mostly
mediated presynaptic inhibition at the calyx–MNTB synapse. Lack of contribution of exocytotic machinery to GABA receptor-mediated presynaptic inhibition DISCUSSION
To examine whether the exocytotic process downstream of Ca2ϩ
Inhibition of calcium currents and transmission by GABA receptor through G-protein
bition, we made simultaneous pre- and postsynaptic recordings
In this study, using paired whole-cell recordings from the brain
and compared the IpCa–EPSC relationship between two condi-
stem giant presynaptic terminal and postsynaptic cell, we have
tions: first after baclofen application and then after reduction of
demonstrated that presynaptic GABAB receptors are linked
[Ca2ϩ]o (Takahashi et al., 1996). When baclofen was applied,
through G-proteins to Ca2ϩ channels, thereby suppressing trans-
EPSCs diminished concomitantly with IpCa (Fig. 8A, i, ii). Sim-
mitter release. The IpCa at the calyx–MNTB synapse is almost
ilarly, when [Ca2ϩ]o was reduced by replacement with [Mg2ϩ]o,
exclusively P-type at the age range examined (Forsythe et al.,
both EPSCs and IpCa were diminished in parallel (Fig. 8A, iii,
1998). It is possible that ␥ complex of heterotrimeric G-protein
iv). When the IpCa–EPSC relations were plotted for data ob-
may interact with the ␣1A subunit, thereby suppressing P-type
tained after baclofen application and after [Ca2ϩ]o reduction, the
Ca2ϩ channel activity (De Waard et al., 1997). Such a membrane-
two relationships largely overlapped with each other (Fig. 8B). At
delimited mechanism is consistent with our finding that the
the seven synapses examined, the slope in the regression lines
magnitude of the baclofen-induced suppression of EPSCs evoked
after baclofen application was similar to that after [Ca2ϩ]o re-
via a presynaptic whole-cell pipette was similar to that of EPSCs
duction (Fig. 8B, inset) (no significant difference in paired t test).
evoked via an extracellular pipette. Thus diffusible intracellular
3144 J. Neurosci., May 1, 1998, 18(9):3138–3146
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
ings from the calyx presynaptic terminals revealed that baclofen
had no effect on the voltage-gated potassium currents or inwardly
rectifying potassium currents. Furthermore, the inward rectifier
channel blockers Ba2ϩ or Csϩ had no effect on the baclofen-
induced inhibition of EPSCs. These results indicate that potas-
sium conductances are not significantly involved in the GABAB
receptor-mediated presynaptic inhibition at this fast excitatory
In our present study, Ba2ϩ had no effect on GABAB receptor-
mediated presynaptic inhibition as reported at other central syn-
apses (Allerton et al., 1989; Lambert at al., 1991; Thompson and
Gahwiler, 1992; Hirata et al., 1995). Although Ba2ϩ was reported
to inhibit the effect of baclofen on monosynaptic IPSCs in hip-
pocampal CA3 cells (Thompson and Gahwiler, 1992), this was
not confirmed in a study using another blocking agent of inward
rectifying potassium channels (Lambert and Wilson, 1993). Fur-
thermore, transgenic mice lacking a GIRK gene exhibited a
normal magnitude of GABAB receptor-mediated presynaptic in-
hibition at hippocampal synapses (Luscher et al., 1997). Thus, so
far there is no direct evidence to indicate an involvement of
potassium conductances in the receptor-mediated presynaptic
inhibition at mammalian central synapses.
At the calyx of Held, an inwardly rectifying potassium current
could be activated by intracellular application of GTP␥S but not
by baclofen. This result may imply that the GABAB receptors and
G-proteins coupled with GIRK are distinct from those coupled
with voltage-gated calcium channels, as proposed previously on
the basis of pharmacological differences between the presynaptic
and postsynaptic effect of baclofen (Dutar and Nicoll, 1988).
However, it is also possible that GIRK is localized outside of the
functional domain of G-proteins coupled with GABAB receptors
Figure 7. Effect of external Ba 2ϩ and Cs ϩ on GABAB receptor-
mediated presynaptic inhibition. EPSCs were evoked in MNTB principal
Baclofen had no effect on presynaptic spike waveform being
cells by extracellular stimulation. A, Baclofen-induced suppression of
consistent with the lack of involvement of potassium conductance
EPSCs was similar before and after Ba 2ϩ application (1 mM). EPSCs
before, during baclofen application (20 M), and after washout are su-
(also see Dittman and Regehr, 1996). Although baclofen sup-
perimposed on top in the absence (left) and presence (right) of Ba 2ϩ.
pressed the presynaptic calcium conductance, this was not appar-
Note the small polysynaptic EPSC component observed at the decay of
ent in the action potential waveform. This might be attributable to
monosynaptic EPSC. B, Baclofen suppressed EPSCs similarly in the
the large potassium conductance masking the calcium conduc-
absence and presence of external Cs ϩ (1 mM). EPSCs before and after
tance. In fact, even after synaptic transmission was abolished by
baclofen application are superimposed on top in the absence (left) and
presence (right) of Cs ϩ. A and B are from different cells.
reducing [Ca2ϩ]o, the presynaptic action potential waveform
remained similar at this (our unpublished observation) and other
messengers, likely to be washed out during whole-cell recording,
synapses (Sabatini and Regehr, 1997).
may not be essentially involved in the baclofen-induced suppres-
sion of EPSCs. IpCa is also suppressed by a metabotropic gluta-
The exocytotic machinery for evoked transmitter release is not affected by GABA receptors
mate receptor (mGluR) agonist (Takahashi et al., 1996). It re-
mains to be seen whether a common G-protein mediates the
The IpCa–EPSC relationship during baclofen application was
presynaptic inhibition by mGluRs and GABA
similar to that during reduction of [Ca2ϩ]o. This suggests that the
exocytotic machinery downstream of Ca2ϩ entry is not involved
Presynaptic potassium channels are not coupled with
in GABAB receptor-mediated presynaptic inhibition at the calyx–
GABA receptor
MNTB synapse, as is the case for mGluR-mediated presynaptic
Presynaptic potassium conductances are thought to be important
inhibition (Takahashi et al., 1996). In the case of baclofen, similar
in the regulation of transmitter release (Augustine, 1990). The
conclusions were made from studies using Ca2ϩ indicators at
receptor-mediated inhibition of a potassium conductance is
hippocampal synapses (Wu and Saggau, 1995; Dittman and
known to enhance synaptic efficacy in invertebrate nervous sys-
Regehr, 1997) (but see Dittman and Regehr, 1996). The direct
tems (Kandel and Schwartz, 1982). At mammalian neuronal so-
involvement of the exocytotic machinery in receptor-mediated
mata, GABAB receptors potentiate transient potassium currents
presynaptic inhibition has been postulated from the observation
(Saint et al., 1990) or activate inwardly rectifying potassium
that baclofen suppressed the frequency of spontaneous miniature
currents through G-protein activation (Andrade et al., 1986;
synaptic currents in a [Ca2ϩ]o-independent manner (Scanziani et
Sodickson and Bean, 1996). It has been postulated that an en-
al., 1992; Rohrbacher et al., 1997). However, differential modu-
hancement of presynaptic potassium currents may underlie
lations of the frequency of miniature events and the amplitude of
GABAB receptor-mediated presynaptic inhibition (Saint et al.,
evoked synaptic responses by various manipulations are well
1990; Thompson and Gahwiler, 1992). However, direct record-
known (Fu and Poo, 1991; Geppert et al., 1994; Cummings et al.,
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3145 Figure 8. Comparison of IpCa–EPSC relationships during baclofen application and [Ca2ϩ]o reduction. Paired recording from a calyx and its target cell. A, Effects of baclofen (20 M; i, ii) and [Ca2ϩ]o reduction (iii, iv) on IpCa (Pre) and EPSCs (Post). IpCa was evoked by 1 msec depolarizing command
pulse from Ϫ70 mV to Ϫ10 mV. Records before and after baclofen application or [Ca2ϩ]o reduction are superimposed on top row. B, Double logarithmic
plot of IpCa–EPSC relation during baclofen application ( filled circles with a dotted regression line) and [Ca 2ϩ]o reduction (open circles with a solidregression line). Data points above 90% in EPSC amplitude were excluded from these plots to minimize constrainment. The slope value was 2.29 for
baclofen and 2.33 for [Ca 2ϩ]o reduction, respectively. Excluding the minimal point from each relationship had no significant effect on the slope values
(2.15 and 2.09, respectively, for baclofen and [Ca2ϩ]o reduction). Inset graph, The slope value of regression lines compared between [Ca2ϩ]o reduction
and baclofen application at seven synapses. No significant difference with p ϭ 0.24 in paired t test. The mean slope value was 1.73 Ϯ 0.17 for baclofen
and 1.77 Ϯ 0.17 for [Ca2ϩ]o reduction, respectively.
1996; Hori et al., 1996). A distinct mechanism may operate in the
receptors at a giant excitatory synapse in rat auditory brainstem slices.
modulation of synchronous and asynchronous transmitter release.
In conclusion, this study has demonstrated G-protein-coupled
Borst JGG, Helmchen F, Sakmann B (1995) Pre- and postsynaptic
whole-cell recordings in the medial nucleus of the trapezoid body of the
modulation of presynaptic Ca2ϩ channels on activation of
rat. J Physiol (Lond) 489:825–840.
GABAB receptor-mediated presynaptic inhibition; neither potas-
Cummings DD, Wilcox KS, Dichter MA (1996) Calcium-dependent
sium channels nor modulation of the exocytotic machinery down-
paired-pulse facilitation of miniature EPSC frequency accompanies
stream of Ca2ϩ influx plays a significant role. On binding a ligand,
depression of EPSCs at hippocampal synapses in culture. J Neurosci
B receptors activate G-proteins and suppress
De Waard M, Liu H, Walker D, Scott VES, Gurnett CA, Campbell KP
Ca2ϩ currents, thereby reducing transmitter release. Given the
(1997) Direct binding of G-protein ␥ complex to voltage-dependent
wide distribution of presynaptic GABAB receptors at synapses
calcium channels. Nature 385:446–450.
throughout the nervous system, this mechanism would be of
Dittman JS, Regehr WG (1996) Contributions of calcium-dependent
and calcium-independent mechanisms to presynaptic inhibition at a
cerebellar synapse. J Neurosci 16:1623–1633.
Dittman JS, Regehr WG (1997) Mechanism and kinetics of heterosyn-
REFERENCES
aptic depression at a cerebellar synapse. J Neurosci 17:9048–9059.
Allerton CA, Boden PR, Hill RG (1989) Actions of the GABA
Dolphin AC, Scott RH (1987) Calcium channel currents and their inhi-
(Ϫ)-baclofen, on neurones in deep dorsal horn of the rat spinal cord in
bition by (Ϫ)-baclofen in rat sensory neurones: modulation by guanine
vitro. Br J Pharmacol 96:29–38.
nucleotides. J Physiol (Lond) 386:1–17.
Andrade R, Malenka RC, Nicoll RA (1986) A G-protein couples sero-
Dutar P, Nicoll RA (1988) Pre- and postsynaptic GABAB receptors in
the hippocampus have different pharmacological properties. Neuron
B receptors to the same channels in hippocampus.
Augustine GJ (1990) Regulation of transmitter release at the squid giant
Forsythe ID (1994) Direct patch recording from identified presynaptic
synapse by presynaptic delayed rectifier potassium current. J Physiol
terminals mediating glutamatergic EPSCs in the rat CNS, in vitro.
Barnes-Davies M, Forsythe ID (1995) Pre- and postsynaptic glutamate
Forsythe ID, Tsujimoto T, Barnes-Davies M, Cuttle MF, Takahashi T
3146 J. Neurosci., May 1, 1998, 18(9):3138–3146
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
(1998) Inactivation of presynaptic calcium current contributes to syn-
Luscher C, Jan LY, Stoffel M, Malenka RC, Nicoll RA (1997) G
aptic depression at a fast central synapse. Neuron, in press.
protein-coupled inwardly rectifying K ϩ channels (GIRKs) mediate
Fu W-M, Poo M-M (1991) ATP potentiates spontaneous transmitter
postsynaptic but not presynaptic transmitter actions in hippocampal
release at developing neuromuscular synapses. Neuron 6:837–843.
Geppert M, Goda Y, Hammer RE, Li C, Rosahl TW, Stevens CF, Sudhof
Mintz IM, Bean BP (1993) GABAB receptor inhibition of P-type Ca2ϩ
TC (1994) Synaptotagmin 1: a major Ca 2ϩ sensor for transmitter
channels in central neurons. Neuron 10:889–898.
release at a central synapse. Cell 79:717–727.
Pfrieger FW, Gottmann K, Lux HD (1994) Kinetics of GABAB
Hagiwara S, Miyazaki S, Rosenthal NP (1976) Potassium current and
receptor-mediated inhibition of calcium currents and excitatory synap-
the effect of cesium on this current during anomalous rectification of
tic transmission in hippocampal neurons in vitro. Neuron 12:97–107.
the egg cell membrane of a starfish. J Gen Physiol 67:621–638.
Rohrbacher J, Jarolimek W, Lewen A, Misgeld U (1997) GABAB
Hagiwara S, Miyazaki S, Moody W, Patlak J (1978) Blocking effects of
receptor-mediated inhibition of spontaneous inhibitory synaptic cur-
barium and hydrogen ions on the potassium current during anomalous
rents in rat midbrain culture. J Physiol (Lond) 500:739–749.
rectification in the starfish egg. J Physiol (Lond) 279:167–185.
Sabatini BL, Regehr WG (1997) Control of neurotransmitter release by
Halliwell JV, Adams PR (1982) Voltage-clamp analysis of muscarinic
presynaptic waveform at the granule cell to Purkinje cell synapse.
excitation in hippocampal neurons. Brain Res 250:71–92.
Hirata K, Ohno-Shosaku T, Sawada S, Yamamoto C (1995) Baclofen
Saint DA, Thomas T, Gage PW (1990) GABAB agonists modulate a
inhibits GABAergic transmission after treatment with type-specific
transient potassium current in cultured mammalian hippocampal neu-
calcium channel blockers in cultured rat hippocampal neurons. Neuro-
Scanziani M, Capogna M, Gahwiler BH, Thompson SM (1992) Presyn-
Hori Y, Endo K, Takahashi T (1996) Long-lasting synaptic facilitation
aptic inhibition of miniature excitatory synaptic currents by baclofen
induced by serotonin in superficial dorsal horn neurones of the rat
and adenosine in the hippocampus. Neuron 9:919–927.
spinal cord. J Physiol (Lond) 492:867–876.
Scholtz KP, Miller RJ (1991) GABAB receptor-mediated inhibition of
Kandel ER, Schwartz JH (1982) Molecular biology of learning: modu-
Ca 2ϩ currents and synaptic transmission in cultured rat hippocampal
lation of transmitter release. Science 218:433–443.
neurones. J Physiol (Lond) 444:669–686.
Kaupmann K, Huggel K, Heid J, Flor PJ, Bischoff S, Mickel SJ,
Sodickson DL, Bean BP (1996) GABAB receptor-activated inwardly rec-
McMaster G, Angust C, Bittiger H, Froestl W, Bettler B (1997) Ex-
tifying potassium current in dissociated hippocampal CA3 neurons.
pression cloning of GABAB receptors uncovers similarity to metabo-
tropic glutamate receptors. Nature 386:239–246.
Takahashi T (1990) Inward rectification in neonatal rat spinal motoneu-
Kubo Y, Reuveny E, Slesinger PA, Jan YN, Jan LY (1993) Primary
rones. J Physiol (Lond) 423:47–62.
structure and functional expression of a rat G-protein-coupled musca-
Takahashi T, Forsythe ID, Tsujimoto T, Barnes-Davies M, Onodera K
rinic potassium channel. Nature 364:802–806.
(1996) Presynaptic calcium current modulation by a metabotropic glu-
Lambert NA, Wilson WA (1993) Discrimination of post- and presynap-
tamate receptor. Science 274:594–597.
tic GABAB receptor-mediated responses by tetrahydroaminoacridine
Thompson SM, Gahwiler BH (1992) Comparison of the actions of bac-
in Area CA3 of the rat hippocampus. J Neurophysiol 69:630–635.
lofen at pre- and postsynaptic receptors in the rat hippocampus in vitro.
Lambert NA, Wilson WA (1996) High-threshold Ca 2ϩ currents in rat
hippocampal interneurones and their selective inhibition by activation
Thompson SM, Capogna M, Scanziani M (1993) Presynaptic inhibition
of GABAB receptors. J Physiol (Lond) 492:115–127.
in the hippocampus. Trends Neurosci 16:222–227.
Lambert NA, Harrison NL, Teyler TJ (1991) Baclofen-induced disinhi-
Wu L-G, Saggau P (1995) GABAB receptor-mediated presynaptic inhi-
bition in area CA1 of rat hippocampus is resistant to extracellular
bition in guinea-pig hippocampus is caused by reduction of presynaptic
Ca 2ϩ influx. J Physiol (Lond) 485:649–657.
FIVE MINUTES WITH . JOHN T. WARD JR. Kentucky Derby-winning trainer John T. WardJr. assumed his new duties as executive direc-Ward, a central Kentucky native and a graduate oftor of the Kentucky Horse Racing Commissionthe University of Kentucky, grew up immersed in thethis spring with an overflowing agenda of is-horse industry as his father, grandfather, and uncle allsues facing the regu
____________________________________________________________________________ Az Internet az egészségügyben Készült a Technológiai Előretekintési Program (TEP) Élettudományokkal és Egészségüggyel folalkozó munkacsoportjának felkérésére Készítette:Dr. Kovács Gábor címzetes egyetemi tanár az MTA doktora Budapest, 1998. November _______________________
 Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3139
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3139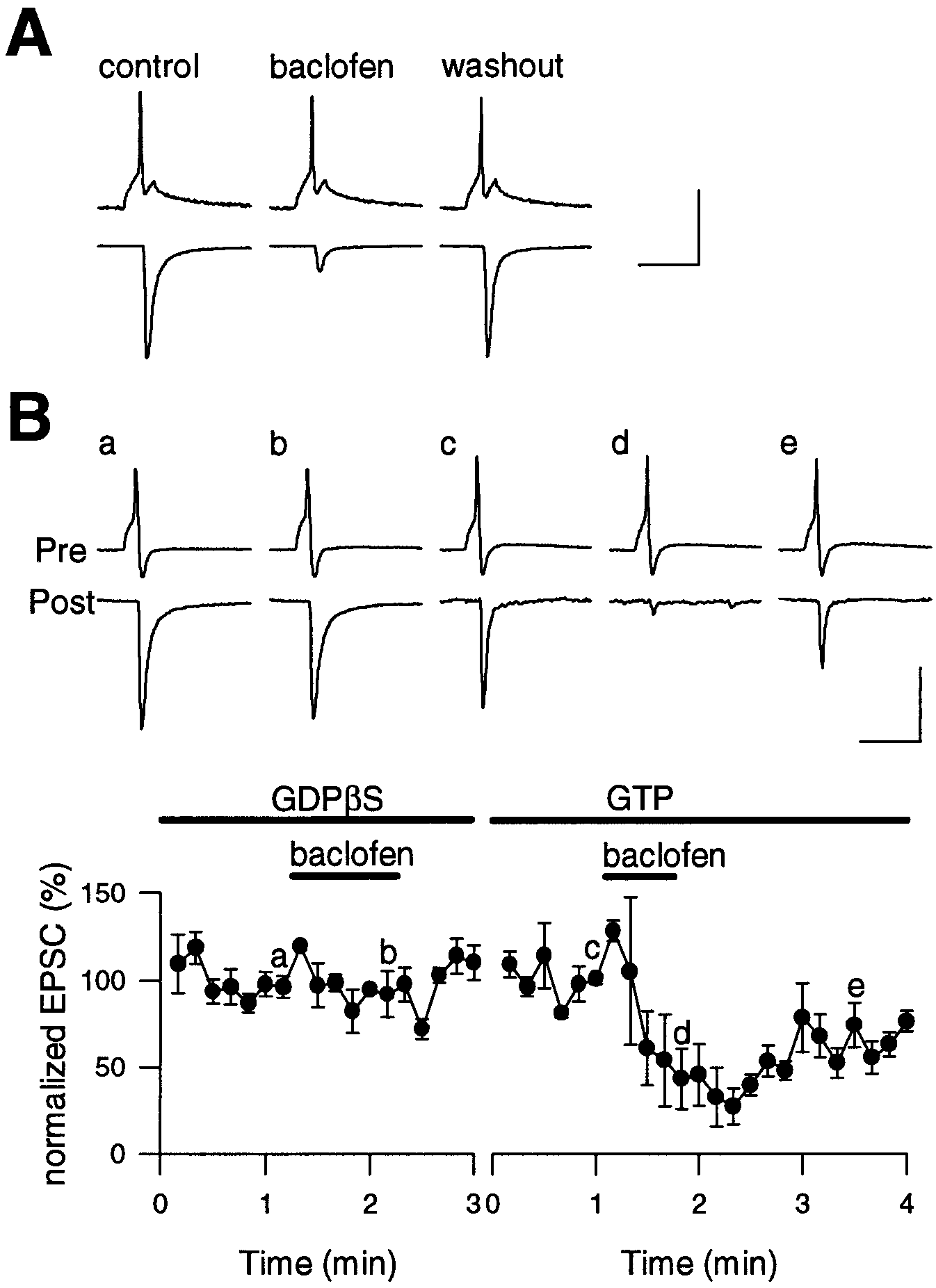 3140 J. Neurosci., May 1, 1998, 18(9):3138–3146
3140 J. Neurosci., May 1, 1998, 18(9):3138–3146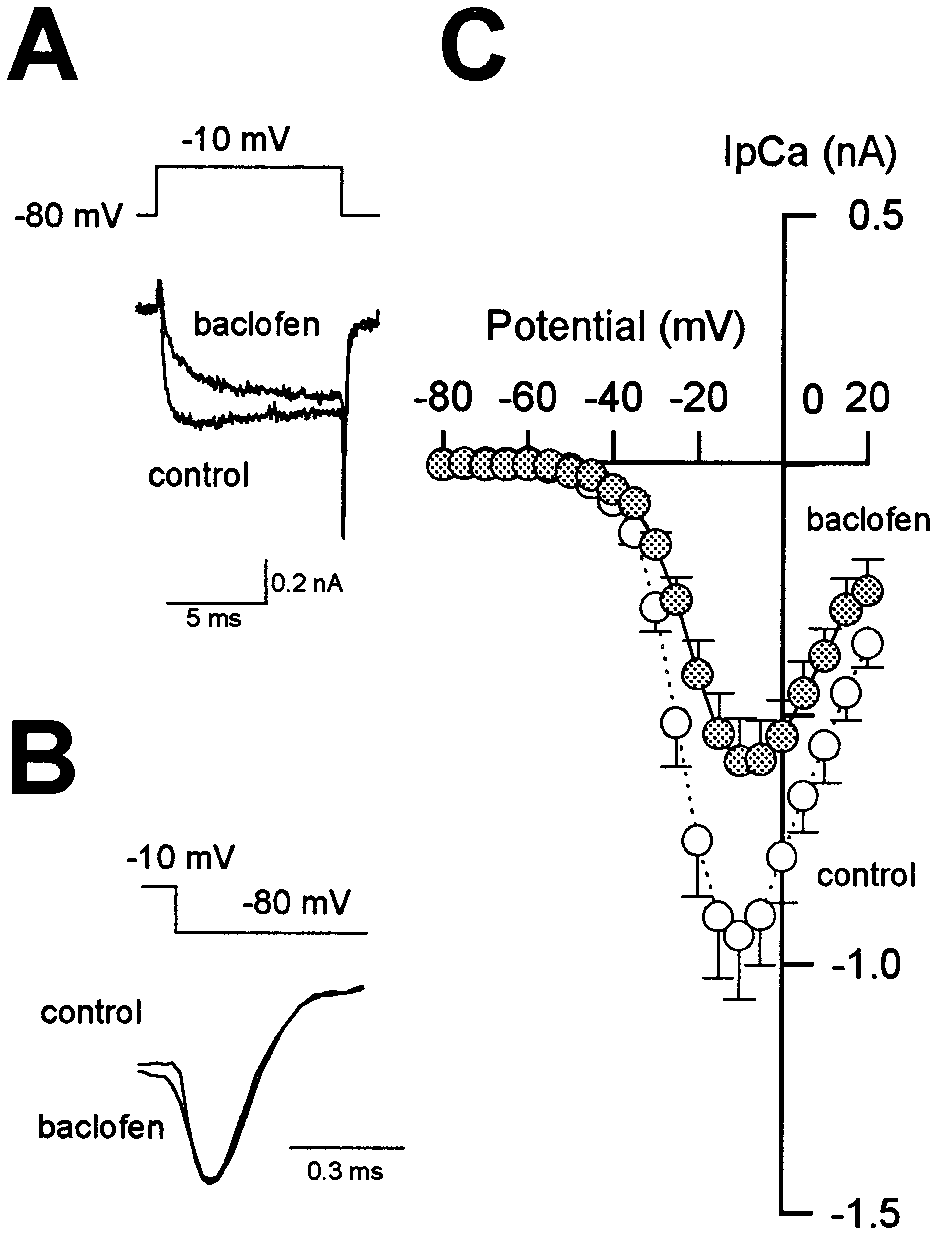 Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3141
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3141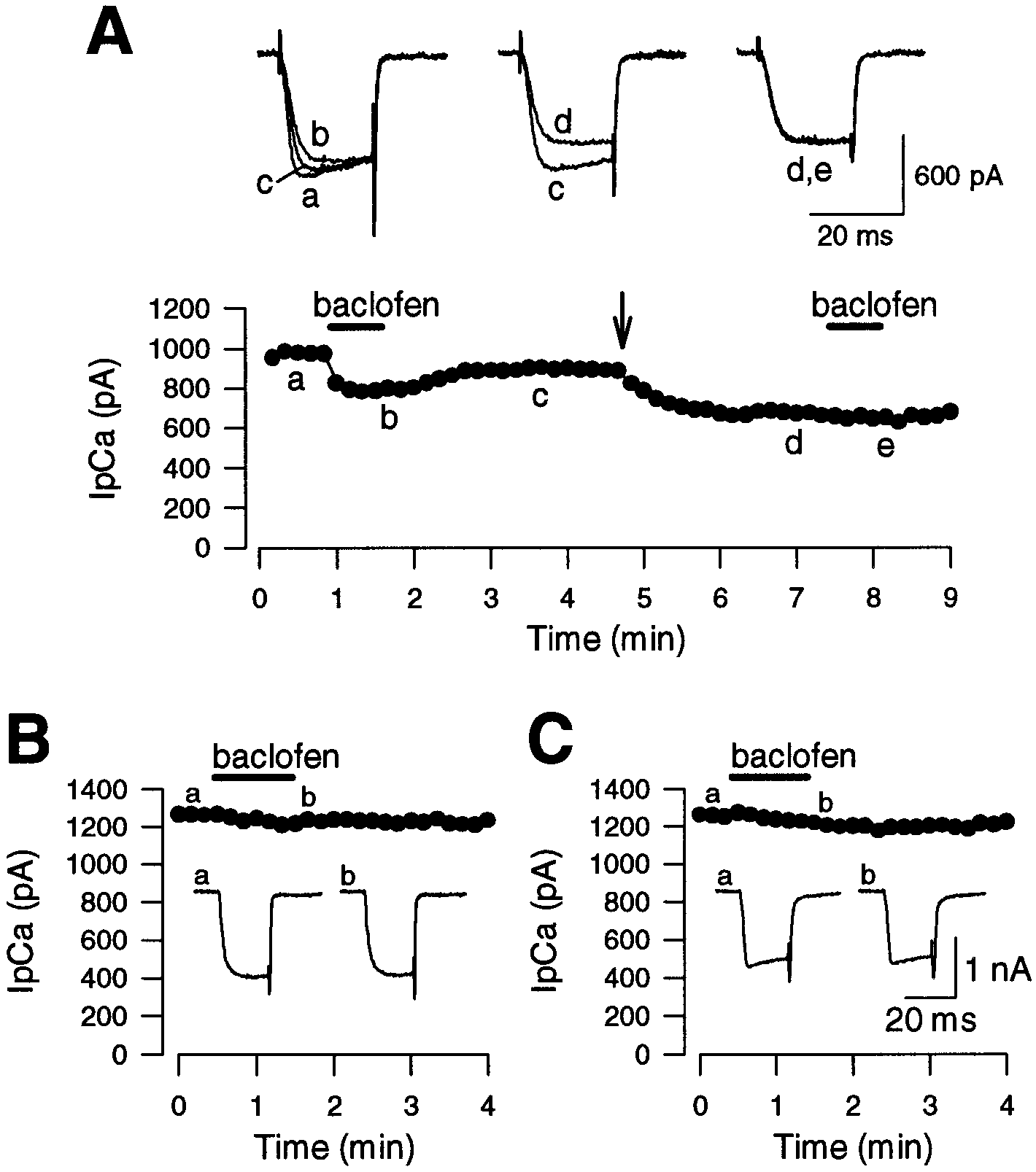
 3142 J. Neurosci., May 1, 1998, 18(9):3138–3146
3142 J. Neurosci., May 1, 1998, 18(9):3138–3146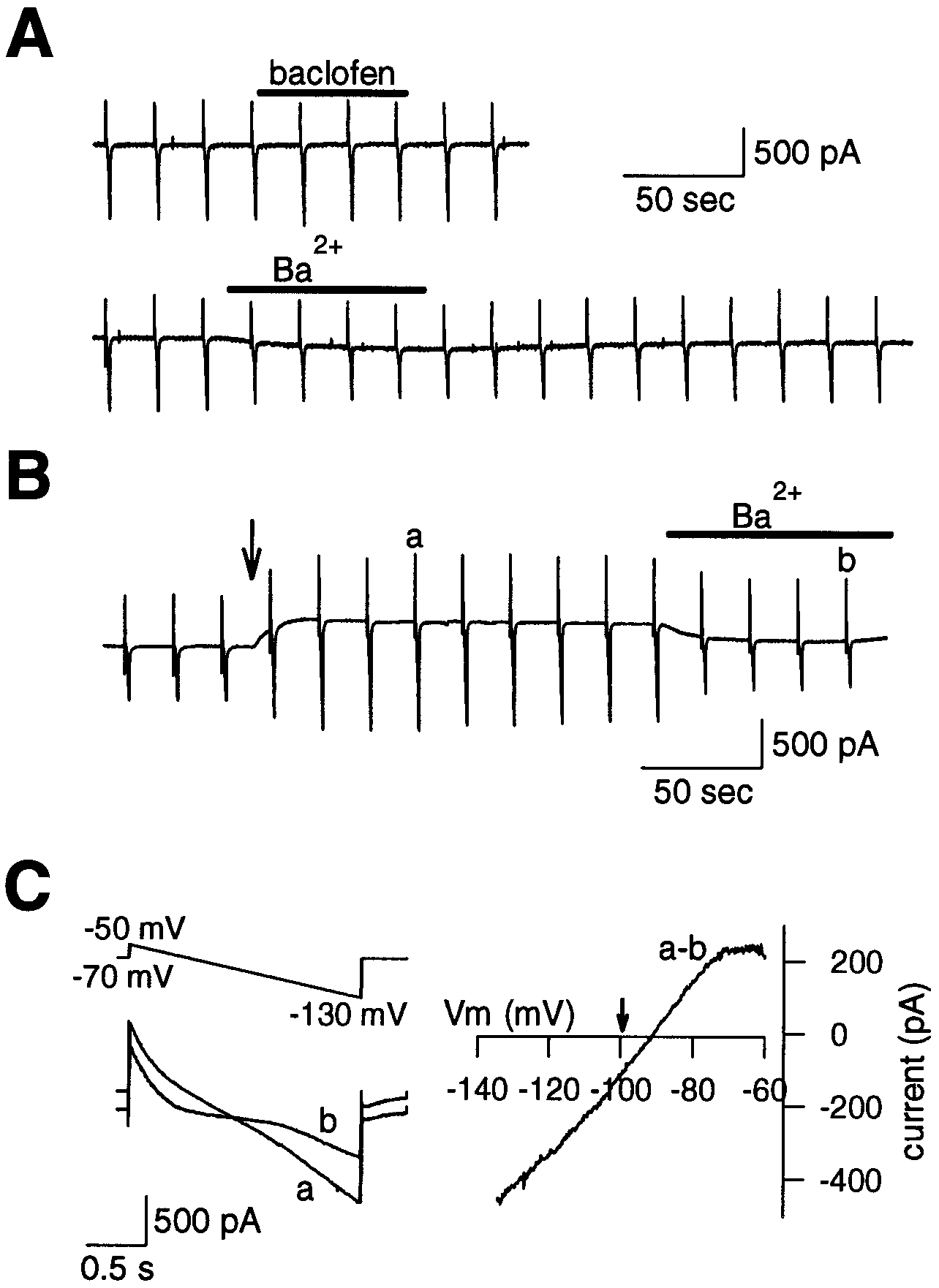 Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3143
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3143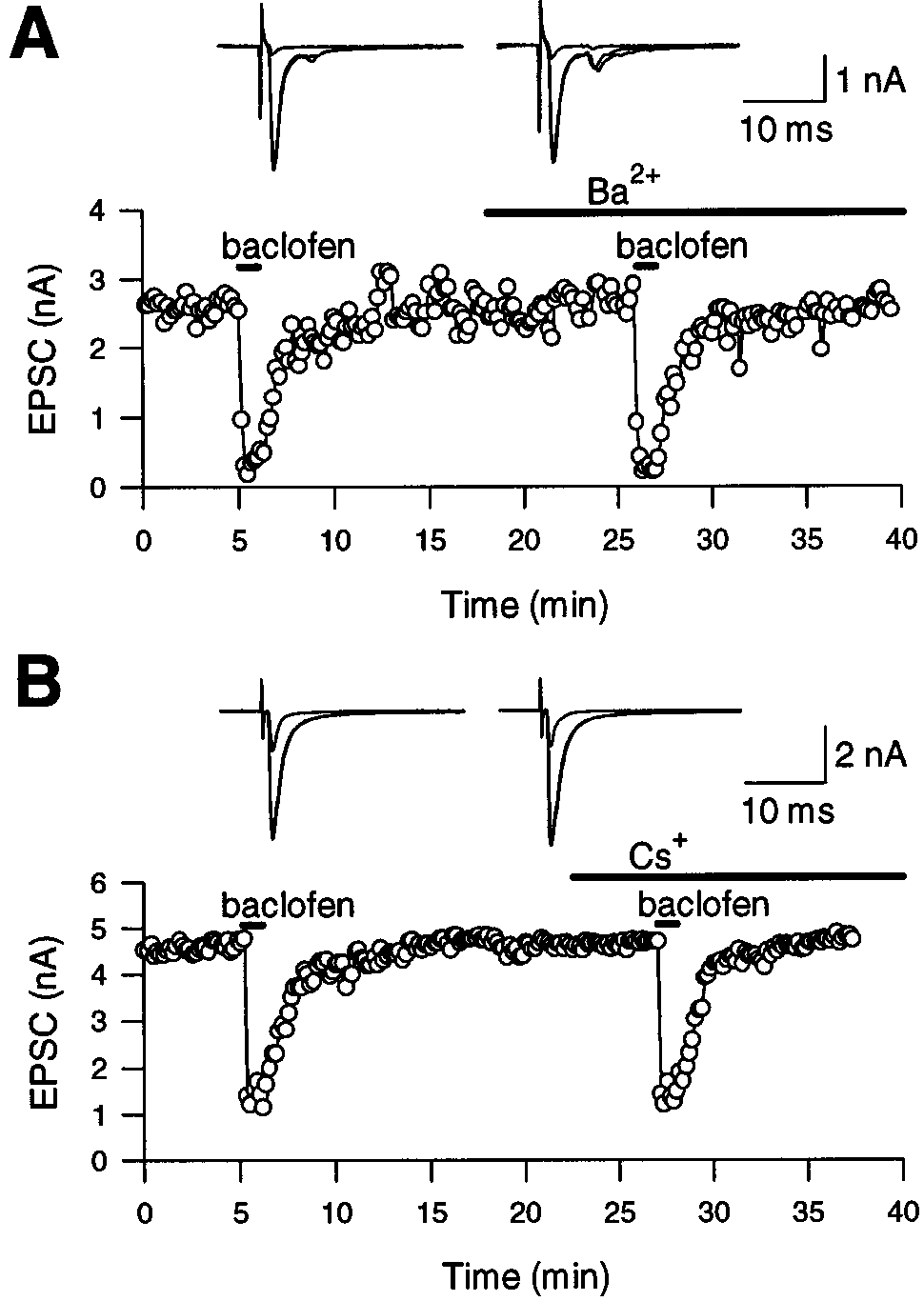 3144 J. Neurosci., May 1, 1998, 18(9):3138–3146
3144 J. Neurosci., May 1, 1998, 18(9):3138–3146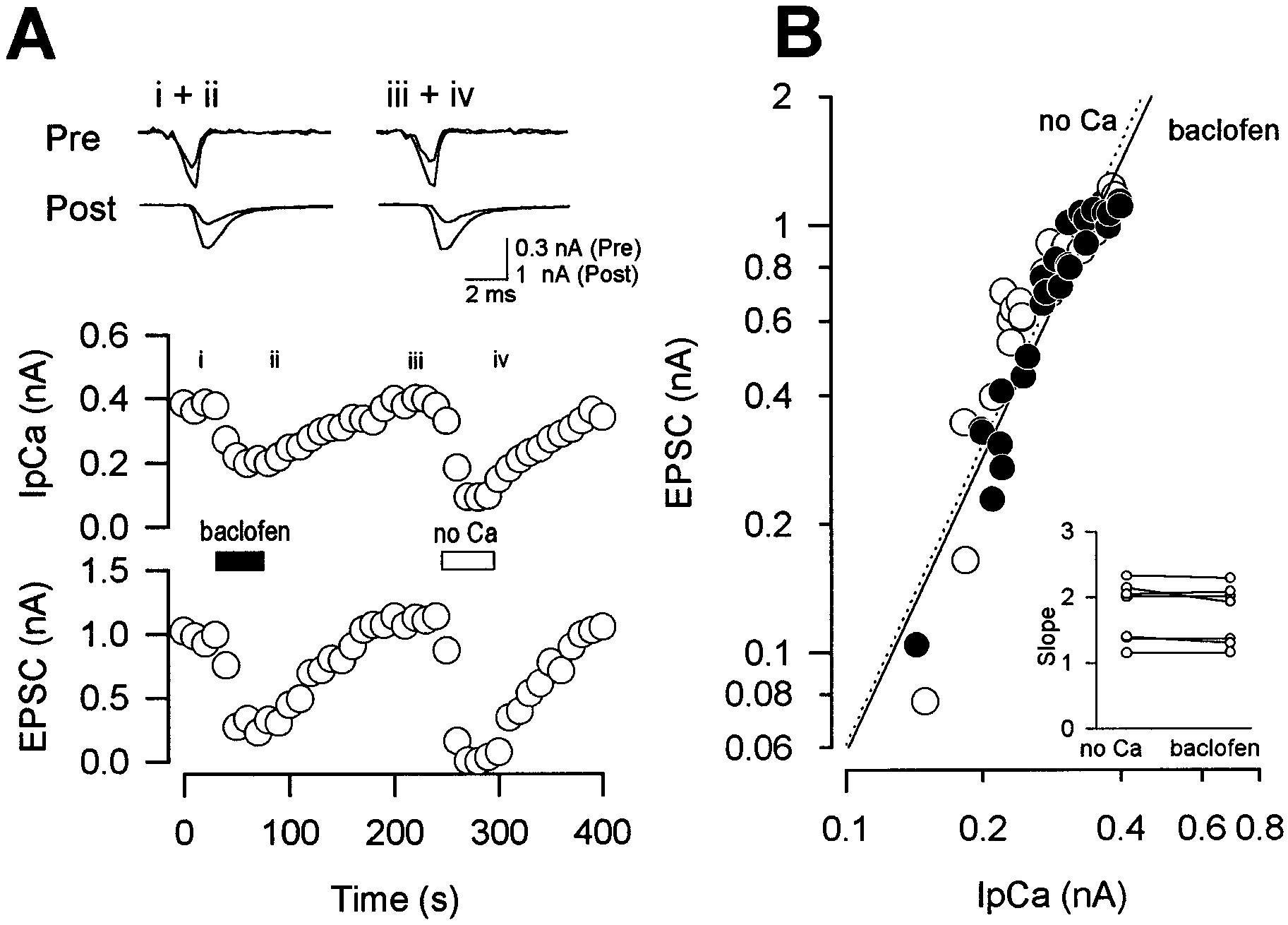 Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3145
Takahashi et al. • Presynaptic Ca Current Modulation by a GABA Receptor
J. Neurosci., May 1, 1998, 18(9):3138–3146 3145