La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Nov jaalas ref 07.indd
Journal of the American Association for Laboratory Animal Science
by the American Association for Laboratory Animal Science
Overview Biologic Effects of Fenbendazole in Rats and Mice: A Review David Villar, Carolyn Cray,* Julia Zaias, and Norman H Altman This review summarizes fi ndings from toxicologic, carcinogenic, immunologic, and metabolic studies on fenbendazole (FBZ). Currently, FBZ is used to treat or prevent pinworm outbreaks in laboratory rodents. Because antiparasitic treatments usually are not part of experimental designs, interactions from the medication on the outcomes of ongoing experiments are a concern. At therapeutic levels, FBZ does not alter the total content of cytochromes P450 but does induce certain hepatic cytochrome P450 isoforms, namely 1A1, 1A2, and 2B1. Although expressed constitutively at low or undetectable levels, these isoforms particularly are known for bioactivating a number of procarcinogens. Lifetime studies in rats have shown that FBZ is not a carcinogen but that it may behave as a tumor promoter when given after certain initiators. Unlike in other animal species, FBZ treatment-associated myelosuppression has not been reported to occur in rodents. The few currently available immunologic studies in mice, including an autoimmune model, have not shown effects on selected immune responses. How- ever, data from other animal species suggest that the ability of B and T lymphocytes to proliferate in the secondary immune response may be suppressed during treatment with FBZ. Abbreviations: EROD, ethoxyresorufi n O-deethylation; FBZ, fenbendazole; GST-P, glutathione S-transferase placental form; MeIQx, 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline; MROD, methoxyresorufi n O-deethylation; MTD, maximal tolerated dose; OXF, oxfenbendazole; ppm, parts per million
Fenbendazole (FBZ) is a broad-spectrum benzimidazole
A review of the treatments available for the eradication of
anthelminth currently approved for use in numerous animal pinworm infestations from laboratory rodent colonies was pub-species, including rodents. Although nematodes, and in particu-
lished recently.53 Of the various agents available, those currently
lar pinworms (for example, Syphacia and Aspicularis spp.), are
used most frequently are avermectins (for example, ivermectin)
the main endoparasites of concern in laboratory rodents, FBZ
and benzimidazoles (for example, FBZ).53 Both drugs are 100%
also is indicated for use in other animal species against a wide
effective by the oral route and typically are delivered in the diet
spectrum of nematodes, tapeworms, fl ukes, and protozoa.9,52
or drinking water. Ivermectin is very safe in adult animals, but
Pinworm infestation in rodent laboratories occurs occasionally,
it can cause toxicosis in young animals due to immaturity of the
and national surveys among large research institutions indicate
blood–brain barrier or in transgenic mice with P-glycoprotein
these incidents are more prevalent than are infections by other
organisms.31 Although the parasites are relatively nonpathogenic,
With regard to the benzimidazole group, FBZ-medicated
their presence may signifi cantly alter the outcome of some labora-
feed has gained widespread use in pinworm prophylaxis and
tory experiments. For example, myelopoiesis and erythropoiesis
treatment protocols because of its large margin of safety and
are known to be increased in pinworm-infected mice and so may
effi cacy. From a practical standpoint, replacing regular feed with
be the sensitivities of bone marrow progenitors to interleukins.7
FBZ-medicated feed adds minimal personnel labor costs and can
In addition, the stimulation of various immune responses, be implemented immediately. In addition, FBZ has adulticidal, including autoimmune responses and elevated production larvicidal, and ovicidal actions.53 Therapeutic levels can be ad-of numerous interleukins, has been linked to infestation with ministered for life without side or toxic effects, as summarized pinworms.1,42 In addition to the multiple effects on the immune
in a report by the World Health Organization.77 However, as
system caused by parasitism, alterations in animal physiology with any other drug, the question arises regarding whether FBZ that would be expected from conditions of mild chronic stress causes physiologic changes that interfere with the outcomes may occur, including changes on neuroendocrine responses, of laboratory experiments. The following sections summarize exploratory behavior, and growth of young animals.41,68,75 Heavy
biologic effects of FBZ in rodents that may be relevant to ex-
infestations may cause overt clinical disease with signs of rectal
perimental protocols in biomedical research.
prolapse, rough hair coats, and general poor body condition.71 These studies highlight the importance of maintaining laboratory
Use of FBZ in Rodents and General Safety
The recommended FBZ therapeutic dosages are usually lower
Received: 20 Jun 2007. Revision requested: 4 Aug 2007. Accepted: 10 Sep 2007.
for livestock species (5 to 10 mg/kg orally once daily for 3 to 5 d)
Division of Comparative Pathology, Miller School of Medicine, University of Miami, Miami, FL.
than for pet animals, including birds and reptiles (20 to 100 mg/
*Corresponding author. Email: ccray@med.miami.edu
kg orally once daily for 3 to 10 d).52 Commercially available med-
Biological effects of fenbendazole in rodents
icated rodent diets contain FBZ at 150 parts per million to reach
Tumor Promoter Studies
a target dosage of 8 to 12 mg/kg daily. In experiments where
As previously mentioned, lifetime studies in mice and rats
actual intake of medicated feed intake by rats was estimated, the
indicate that FBZ itself is not a carcinogen. Histologic changes
mean daily consumption rate for male rats was 8.4 mg/kg and
consisting of hepatocellular hypertrophy, bile duct proliferation,
that for female rats was 11.5 mg/kg.11 These feed concentrations,
hyperplasia, and vacuolation occur in the livers of rats given
given in alternating weeks, effectively eradicated pinworms in
at least 45 mg/kg.77 The WHO joint expert committee viewed
rat and mouse colonies.4,11,23,27,28 Although colony infestations
these changes as an adaptive response to toxicity; the question
have been eliminated without environmental decontamination
of whether FBZ could serve as a tumor promoter remained
or changes in husbandry practices, re-emergence of pinworms
unresolved and was addressed later.65,77 Dosages of 45 mg/
have occurred when apparently adequate surveillance programs
kg or greater in rats likely achieved toxicity, and the histologic
features of proliferation were secondary to chronic insult to the
A World Health Organization Joint Expert Committee on liver. Cells in chronically injured tissues are exposed continu-
Food Additives compiled a comprehensive report on FBZ that
ously to endogenous mitogens (for example, growth factors),
included an evaluation of unpublished acute toxicity studies which ultimately promote cancer development through clonal
and long-term toxicologic data in rodents.19-21,60,61,77 FBZ can be
considered a nontoxic drug because in rodents, the dose lethal
Carcinogenesis is a multistage continuous and dynamic pro-
to 50% of the tested population exceeds 10 g/kg (a dose 1000
cess that is conceptually divided experimentally into the stages
times the therapeutic level). In a short-term (14 d) toxicity study
of initiation, promotion and progression. Therefore, even when
using Sprague–Dawley rats (weight, 180 g), doses equal to or
not a complete carcinogen itself, a compound can contribute to
greater than 50 mg/kg slowed body weight gains.64 Although
cancer susceptibility by promoting proliferation of previously
overt clinical signs were not noted at 500 and 3000 mg/kg, these
initiated cells. For example, compounds (such as dioxin), which
doses caused histopathologic degenerative changes in the liver
cause no DNA damage and are negative by the Ames test, are
and kidney.64 In a 90-d subchronic study, doses of 1600 mg/kg
potent tumor promoters.40 Tumor promoters typically act over
for 60 d followed by 2500 mg/kg for another 30 d did not cause
time, and what is initially a reversible lesion eventually develops
clinical signs or pathologic effects.77 However, the actual data for
into cancer after prolonged exposures to a promoter agent.
that study were not presented, and because the report is incon-
To resolve whether FBZ acts as a tumor promoter, a medium-
sistent with the toxicity data from short-term (14 d) studies or
term liver bioassay known to be a reliable test for the detection of
even those with data from longer exposures, whether pathologic
carcinogens as well as promoters of hepatocarcinogenesis, was
changes (for example, increased liver weight and hepatocellular
applied to FBZ and its metabolite oxfenbendazole (OXF).43,63,65
hypertrophy) were, in fact, nonexistent is unclear.64
The assay was a 2-stage liver carcinogenesis model in which
When lifetime toxicity–carcinogenicity exposures were started
rats were initiated with a single in vivo dose of the genotoxic
in utero and terminated at week 123 in Sprague–Dawley rats, hepatocarcinogen diethylnitrosamine; 1 wk later they began
doses of 135 mg/kg were associated with reduced body weight
receiving a diet containing FBZ at 0, 70, 200, 600, 1800, and
at euthanasia, decreased survival (25% versus 35%), and slightly
3600 ppm for 8 wk. One of the biomarkers to assess promo-
increased incidence of hepatocellular carcinoma.77 However, tion (decrease in gap-junctional intercellular communication
because the maximal tolerated dose (MTD) had been exceeded,
connexin 32) appeared to indicate a positive effect at doses
the results from this group could not be used for assessment of
equal to or greater than 70 ppm, the other marker (glutathione
carcinogenesis. According to current Food and Drug Administra-
S-transferase placental form, GST-P) was only positive at doses
tion guidelines, the main criteria for setting an MTD is that the
of or exceeding 1800 ppm.65 Given these results, the authors
animals remain in good clinical condition and do not lose or fail
proposed that FBZ has liver tumor-promoting activity similar
to gain body weight to an extent greater than 10% of similarly
aged controls.54 This defi nition allows subtle biochemical and
Because there are no standard criteria for classifying a com-
cellular end points, such as induced mitogenesis, at the MTD. In
pound as a promoter, the experimental conditions and type of
lifetime studies, an FBZ dose of 45 mg/kg, which approximates
biomarkers used are an important source of variation for this
the MTD, caused morphologic changes of hepatocellular hyper-
type of study. Of the numerous biochemical markers for early
trophy and hyperplasia.77 A pathology working group viewed
detection of preneoplastic cells, the number and areas of GST-P–
the histopathologic changes as an adaptive response to toxicity
positive liver foci undoubtedly have been the most widely used
unrelated to the formation of hepatic neoplasms. The group endpoints, because they correspond well with the incidence of
reached 2 other conclusions: 1) the lifetime no-observed adverse
hepatocellular carcinomas in long-term in vivo assays.51,63 GST-P
effect level for maternal and reproductive toxicity in rats was set
is an enzyme strongly expressed in so-called initiated cells, but
at 15 mg/kg daily, and 2) doses higher than 15 mg/kg increase
not normal hepatocytes, during the early stage of chemically
the incidence of hepatocellular altered foci and hypertrophy. induced hepatocarcinogenesis.58 By considering this assay as
Recent fi ndings that administration of a 45 mg/kg dosage to rats
the hallmark to identify preneoplastic hepatic foci, we can state
for only 2 mo increased relative liver weights, caused periportal
that FBZ seems to act in a manner similar to tumor promoters,
hepatocellular hypertrophy, and increased mitotic activity in rats
although at clinically toxic doses (that is, doses exceeding the
led to the conclusion that this dosage triggers cell proliferation.65
MTD). Therefore, the results of GST-P assays would not be
From the cited studies, the 45 mg/kg dosage likely was close noteworthy in terms of risk at therapeutic levels of FBZ.
to the MTD for carcinogenicity studies with rats, although no
This is not the case for the inhibition of connexin 32 by FBZ,
particular dosage has been accepted as such.
which occurs at much lower and noncytotoxic levels within
Mice appear to be less sensitive to FBZ than are rats. However,
the therapeutic range for FBZ (70 to 200 ppm).65 Decreased ex-
little species-specifi c information is available. A 2-y carcinoge-
pression of connexin 32, the predominant hepatic gap junction
nicity study with doses as high as 405 mg/kg daily did not show
protein, is shared by numerous tumor-promoter agents that
an increase in tumor incidence.77 The no-observed adverse effect
exert their promoting activity through different mechanisms.35,73
level for mice was set at 135 mg/kg daily.77
However and unlike the GST-P endpoint, the inhibition of
Vol 46, No 6Journal of the American Association for Laboratory Animal ScienceNovember 2007
connexin 32 is a necessary but insuffi cient factor to consider a compound as a tumor promoter. Two things are necessary for an initiated cell to proliferate: a) an intracellular signal from the chemical to block contact inhibition and transfer of signals from cell to cell via gap junctions, and b) an intracellular signal to proliferate. Disruption of gap junctions does not necessarily imply that the chemical induces a mitogenic signal.
A previous study looked at another potential biomarker of tu-
mor promotion: the induction of certain cytochromes P450 such as CYP2B1.65 These are discussed later in the text. Nevertheless, that a compound exerts an inducing effect on cytochrome P450s may not necessarily imply a promoter or mitogenic action; and so, induction should not be considered a reliable endpoint of promoter activity.37 This point is illustrated by the induction of CYP1A2 by FBZ in relation to a known dietary carcinogen, MeIQx (2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline). MeIQx is an heterocyclic amine thought to be metabolically bioactivated to genotoxic intermediates in the liver by CYP1A2.36,74 Combining FBZ (600 ppm in the diet) with MeIQx did not enhance MeIQx-induced hepatocarcinogenesis, despite the fact that FBZ caused a 2.3-fold increase in CYP1A2 levels.70 In this case, only GST-P positive liver cell foci were used to assess carcinogenicity.
When the same 2-stage carcinogenesis model was applied
to OXF, which is the primaryin vivo metabolite of FBZ, lower doses of OXF (10 and 100 ppm) had greater effects at inducing the same cytochromes and affecting the same assays (connexin 32 and GST-P) used as biomarkers of preneoplastic lesions.43 The higher potency of OXF suggests that many of the in vivo effects of FBZ likely were caused through this metabolite.
In conclusion, the medium-term liver bioassay in rats showed
that FBZ may act as a promoter when combined with certain
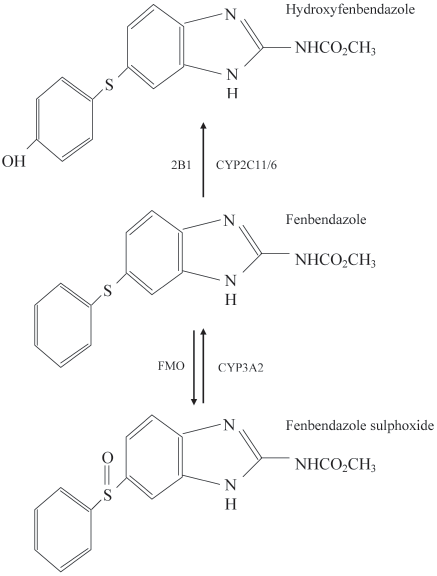
Figure 1. Initial FBZ biotransformation and key cytochrome isoforms
genotoxic chemicals like diethylnitrosamine but not with other
implicated in rats. Total metabolite production toward FBZ sulphox-
compounds, such as MeIQx. However, because no effects were
ide (also called oxfenbendazole, OXF) and hydroxyfenbendazole is
seen in uninitiated groups, these studies also support previous
nearly identical in rats.46,67 These are the 2 main metabolites detected
evidence from long-term carcinogenicity experiments that FBZ
in plasma and primarily are eliminated through bile into feces. Notice
that fenbendazole and OXF are metabolically interconvertible. FMO, fl avin monooxygenase system; CYP, cytochromes.
Effect on Cytochromes P450
The widespread use of FBZ in veterinary medicine has tive concentrations of the common P450 enzymes in rat hepatic
prompted numerous studies on its effect on hepatic biotrans-
formation enzymes. FBZ is biotransformed largely by hepatic
FBZ induces 2 members of the highly conserved 1A subfam-
microsomal P450 and to a lesser extent by fl avin monooxygenase ily (1A1 and 1A2) as well as cytochrome 2B1 in rats.2,65 These enzyme systems (Figure 1).46,66,67 Consequently, any changes in
isoforms are constitutively expressed at very low or virtually
the activity of these enzymes may alter the pharmacokinetics
undetectable levels (Tables 1 and 2), which could explain the lack
of FBZ and other coadministered xenobiotics and divert their
of effect on total P450 contents and associated enzyme activities
normal biotransforming pathways. In fact, goats pretreated in the early studies. The CYP1A1 and 1A2 protein levels in rat with the nonspecifi c P450 inhibitor piperonyl butoxide showed
hepatocytes incubated with different concentrations of FBZ for
more than 3-fold increases in the relative bioavailability (that is,
48 to 72 h increased 8-fold and 7-fold, respectively, along with
area under the curve) of FBZ and its primary metabolite OXF.6
a 2- to 3-fold increase in the levels of the corresponding func-
Interestingly, coadministration of both drugs greatly potentiated
tional markers, ethoxyresorufi n O-deethylation (EROD) and
their antinematodal activity, and this potentiation was attributed
methoxyresorufi n O-demethylation (MROD).2 Rats dosed with
to the extended pharmacokinetic profi le of FBZ.6
200 ppm (that is, 1.3 times therapeutic levels) for 8 wk and eu-
Early studies in rodents showed that FBZ did not exert either
thanized 1 wk later had a 3-fold induction of CYP1A2, whereas
positive or negative effects on total microsomal P450, even at
CYP1A1 remained undetectable.65 However, the magnitude of
100 mg/kg daily for 15 d (that is, at 10 times the therapeutic this induction caused by FBZ can be considered minor when level).12,44 No effect was also found in other microsomal constitu-
compared to classic inducers like 3-methylcholanthrene and
ents, including NADPH cytochrome c reductase, cytochrome β-naphthofl avone, for which increases of several hundredfold b5, and glutathione S-transferase. These studies were conducted
are typical after 24 h of treatment (Table 2). Studies in primary
before the advent of more recent molecular biology techniques
cultures of rabbit hepatocytes have shown that, at least for
that have classifi ed the P450 system into families, subfamilies
CYP1A1, the mechanism of FBZ induction involves transcrip-
and specifi c isoforms; so far, at least 93 functional cytochrome
genes have been sequenced in mouse liver, with 82 members
From a practical standpoint, CYP1A1 and 1A2 have received
belonging to the 4 major drug-metabolizing families.39 Table 1
much attention because they are well known for activating a
provides further information illustrating the content and rela-
number of procarcinogens, such as aromatic amines present in
Biological effects of fenbendazole in rodents
Table 1. Concentrations of P450 enzymes in rat hepatic microsomes
Data deduced from references 47 (study 1) and 24 (study 2). aDetermined spectrally. bSum of enzymes determined immunochemically. Note that 30.5% (study 1) and 49% (study 2) of the other CYP isoforms were not determined.
organic pyrolysis products and polycyclic aromatic hydrocar-
Bone Marrow Effects
bons present in tobacco smoke and charcoal-broiled meat.62
In recent years, clinical observations of myelosuppression
Furthermore, a recent critical review provides strong evidence
associated with FBZ treatment have been documented for
that the chemoprotective effect of numerous fl avonoids pres-
various animal species including porcupines, canines, pigeons
ent in fruits, vegetables, and plant beverages is, among other
and doves, and tortoises; however, we found no similar re-
mechanisms, through inhibition of the metabolic activation ports for rodents.17,22,26,48,76 A common denominator in these
of procarcinogens by cytochromes P450 1A1 and 1A2.45 The cases of myelosuppression was the diagnosis of bone marrow
cited review also describes in vivo studies showing that some
hypoplasia within a few days of initiating FBZ treatment. The
flavonoids can suppress the tumor formation induced by dosages given were all in the upper end of the recommended
polycyclic aromatic hydrocarbons and other carcinogens in range (50 to 100 mg/kg for several days) and at least for birds,
experimental animals.45 The wide application of genetic en-
a possible dose relationship was mentioned (that is, higher
gineering and polymerase chain reaction techniques to better
morbidity and mortality in birds given higher doses), together
defi ne the roles of specifi c cytochrome isoforms have revealed
with a greater occurrence in columbiform birds.22,26 However,
that overexpression of the CYP1A1 gene is accompanied by whether myelosuppression develops seems to be unpredictable,
alternative splicing variants of the enzyme that are expressed
and no animal models are available to reproduce the condi-
in compartments other than the endoplasmic reticulum (that tion, suggesting an immune-mediated mechanism of action
is, the nucleus) and facilitates the neoplastic transformation of
or an idiosyncratic reaction in extremely sensitive animals.
normal cells exposed to procarcinogens.38
An idiosyncratic reaction is possible because FBZ also inhibits
Because liver microsomes from rodents have low expression
mammalian microtubule assembly and blocks mitosis of hu-
of CYP1A2 and virtually undetectable levels of CYP1A1 and man lymphocytes at metaphase.13,25 A sensitization reaction
CYP2B1, their overall contribution to total biotransformation that involves the immune system and does not have a dose–
of most xenobiotics is probably small unless they became response relationship is also feasible because very high doses
induced.14,65 Of the 3 cytochromes that have been so far shown
of FBZ that result in toxicosis affect organs (for example, liver,
to be induced by FBZ, only CYP2B1 appears to participate in
kidney) other than the hematopoietic system. The selective
the fi rst oxidation step for FBZ (Figure 1). In rats, the fi rst 2
toxicity of benzimidazole anthelmintics toward nematodes is
oxidations occur through CYP3A and the fl avin-monooxygenase assumed to derive from the greater susceptibility of parasitic β
system, and CYP2C6/11 and CYP2B1 are involved in the conver-
tubulins (compared with that of their mammalian counterparts)
sion to the 4’-hydroxyl metabolite (Figure 1).46,67 By inducing
to inhibition of polymerization. This susceptibility appears to
CYP2B1, FBZ may hasten the oxidation to the FBZ-OH metabo-
correlate well with the affi nity of the benzimidazole drugs for
lite and alter its own bioavailability; this scenario remains to be
binding tubulin, to the extent that determining key amino acid
studied. Similarly, drug interactions from metabolism through
residues in the structure of β tubulin can be used to predict
these inducible FBZ isoforms may be discovered in the future.
resistance to benzimidazole drugs.33,57 Rodents have been used
For example, a 40% lower plasma concentration and faster to understand the mechanisms of drug-induced bone marrow
clearance of propanolol have largely been attributed to a 1.5- to
failure for some compounds such as benzene and chloram-
2-fold induction of CYP1A2 by ginkgo herbal extracts, although
phenicol; however, they may not be suitable animal models
the involvement of other isoforms was not ruled out.79
for FBZ because no myelosuppresive effects in rodents have
In unpublished observations with mice, we found that FBZ
is converted rapidly to OXF when injected intravenously at a dosage of 10 mg/kg. However, the rates of conversion dif-
Effects on the Immune System
fered markedly among animals, with concentrations of OXF
Immunologic function is a critically important variable that
measuring 25% to 400% of those attained for FBZ at 18 to 30 underlies most, if not all, experimental protocols. Several studies
min after injection. Therefore, marked differences in expression
have examined whether FBZ exerts any immunomodulatory
levels of the cytochromes involved in the sulfoxidation of FBZ
effects on the immune system.8,15,16,49,55 Some of these studies,
likely largely account for the extreme interanimal variation in
together with the effects of other anthelmintics, have been re-
Vol 46, No 6Journal of the American Association for Laboratory Animal ScienceNovember 2007
Table 2. Induction of liver P450 isoenzymes (CYP1A1 and 1A2) and the corresponding dealquilation assays (EROD and MROD) by FBZ,
3-methylcholanthrene (3MC), polychlorinated biphenyls (PCB), β-naphthofl avone (βNF), and phenobarbital (PB) in rats
(pmol/mg protein) (pmol/mg protein) (pmol/mg protein)
Control animals received an intraperitoneal injection of corn oil or were untreated; experimental rats were given 3MC (40 mg/kg) or βNF (100 mg/kg) intraperitoneally for 3 d and were euthanized 24 h after the last treatment, one injection of PCB (Aroclor 1254; 500 mg/kg) intraperitone-ally and were euthanized after 5 d, or PB (80 mg/kg) intraperitoneally for 4 d and euthanized 24 h after the last treatment. FBZ (200 ppm) was provided for 8 wk in the diet of diethylnitrosamine-initiated rats. EROD and MROD assays were done with a substrate concentration of 50 μM. The total cytochrome P450 content in the liver microsomes of control rats ranges between 400 and 1000 pmol/mg protein (Table 1).
viewed recently.56 Both stimulatory and suppressive effects on
most noticeable effects occurred after a second challenge to FBZ
different components of the immune system have been reported,
also suggests an immune response in which the drug behaves
adding to the diffi culties of interpreting their physiologic impli-
as a hapten during the initial exposure.
cations. Nevertheless, with the use of genetically modifi ed mice,
These studies in sheep prompted similar experiments in
it may now be possible to relate changes in specifi c components
rodents, which are underway in our laboratory. Recent work
of the system to their physiologic implications to the animal. One
has demonstrated that aged (22-mo-old) BALB/c mice on FBZ
study looked at the effects of FBZ on a mouse model (nonobese
treatment regimens have less B cell proliferation in response to
diabetic, NOD), where T cells become autoreactive against pan-
mitogens than do young (3- to 4-mo-old) BALB/c mice.78 Ad-
creatic islet antigens, thus causing type I diabetes.16 This study
ditional experiments to assess other aspects of the in vitro and
found that exposure of NOD mice to a FBZ-medicated diet for
in vivo immune responses during FBZ treatment are ongoing.
23 wk did not alter the incidence or onset of diabetes compared
In conclusion, there are confl icting reports on whether FBZ
with that of the control group. Furthermore, no changes were
exerts immunomodulatory actions. Although most studies
seen in lymphocytes subpopulations (CD4:CD8) or T lympho-
have shown no effect of FBZ on selected immune responses, the
cyte proliferative responses to Con A. Experiments in common
question of whether FBZ suppresses lymphocyte proliferation
strains of mice (BALB/CByJ, C57BL/6J) fed 100-ppm FBZ diets
remains controversial. Because the immune system is under con-
for 2 wk found no effect on a number of specifi c immune re-
tinuous self-regulation to balance the intensity and specifi city of
sponses: ability to generate helper T cells, allospecifi c cytolytic
its responses, any drug that suppresses lymphocyte proliferation
T cells, priming of pre-killer cells, and production of specifi c
may affect multiple responses such as allergy, autoimmunity,
antibodies against an infl uenza virus.55 A recent retrospective
graft rejections in transplants, and antibody formation.
study reported FBZ did modulate the infl ammatory process in F344 rats.29 During a study period, the rats were treated with
Reproduction, Teratologic, and Behavioral
FBZ as part of a colony management directive. The investiga-tors found that the experimental rats, which were injected with
LPS as part of the study, demonstrated increased weight loss,
Effects on reproductive performance and offspring produc-
microglial activation, and loss of astrocytes.
tion can have dire consequences for researchers, especially when
Extensive studies highlighting the effects of FBZ on immune
dealing with rare transgenic strains. Therefore, the reproduc-
system function have been performed in sheep. These studies
tive, teratogenic, and behavioral effects of FBZ are critically
were conducted in nonparasitized lambs and examined multiple
important to understand. Reproductive studies conducted over
immune responses after the administration of FBZ or OFX.8,49,69
3-generation SD rats at doses of at least 5 mg/kg concluded that
Briefl y, 6-mo-old lambs were drenched with a single dose of dosages of 45 mg/kg or greater caused reduced fertility and se-FBZ or OXF on days 0 and 28, and 1 d after each drench, they
vere signs of toxicosis in pups (for example, decreased survival
were injected with human erythrocytes and ovalbumin. The indices, decreased body weights at birth, slower lactational assessment of the immune system included T cell and B cell growth, and so forth).77 Because the lower dosages of 5 and 15 proliferation assays and antibody responses against the human
mg/kg did not cause signifi cant alterations, the no-observed-
erythrocytes and ovalbumin antigen injected. These studies effect limit for reproductive effects was set at 15 mg/kg daily.77 showed that for some sampling dates, both B and T lympho-
A recent report conducted from retrospective breeding records
cytes collected after the fi rst and particularly after the second
noted an association between litter size (that is, fecundity) and
administration of FBZ or OXF had lower stimulation indices. In
FBZ treatment in rats.32 The rats that were given FBZ feed on
addition, antibody responses were depressed after the second
a intermittent or continuous basis for as long as 7 wk had 3 to
antigen injection. The studies concluded that FBZ and OXF may
4 fewer pups per litter than did nonmedicated animals. No
affect the general ability of circulating lymphocytes to divide,
other endpoints of reproductive toxicity were evaluated (for
particularly those in the secondary immune response. These example, weaning and lactation indices, fertility, stillbirths), and fi ndings are consistent with earlier reports demonstrating the
several factors could have infl uenced the interpretation of the
ability of benzimidazole drugs, including FBZ, to block mitosis
results, including the greater age of the dams on the medicated
of human lymphocytes in culture.25 However, the fact that the
diet and the differing nutrient composition of 2 diets. In the same study, no effect on litter size was observed in genetically
Biological effects of fenbendazole in rodents
Table 3. Biologic effects of FBZ at therapeutic levels (150 ppm; 8 to 12 mg/kg daily) in rodents and other species
connexin 32 after initiation with diethylnitrosamine
epilepsy-prone rats (GEPR, substrain 9) that received a similar
fecundity, there was no effect on body weight gain in pups. Two
additional studies also reported the lack of behavioral effects in
No evidence of teratogenicity was seen at levels as high as rats on FBZ-medicated diets.34,72 In those studies, standard tasks
2500 mg/kg in rats; mice were not studied.77 In another study,
included food search, drinking behavior, and lick rates.
the offspring of SD rats continuously exposed to therapeutic levels of FBZ were examined in a variety of behavioral terato-
Conclusions
logic paradigms.5 Of 5 behaviors examined, 2 (negative geotaxis,
A review of the literature has shown that FBZ at therapeutic
digging maze performance) were unaffected, and 3 (delayed levels will not cause any toxic effects but, like any other drug,
righting refl ex, Morris water maze, and running wheel) showed
it has some physiologic actions that potentially can change the
subtle or minor alterations in performance. However, because
outcome of laboratory experiments (Table 3). Although long-
the effects were subtle, their biologic relevance was question-
term carcinogenicity experiments have proven that FBZ itself is
able, and the overall conclusion was that FBZ had minimal not a complete carcinogen, FBZ given at therapeutic levels after
consequences on behavioral and developmental studies. Al-
certain genotoxic initiators like diethylnitrosamine did inhibit
though the cited study did not report the litter size to assess for
gap junction intercellular communication, which is a common
Vol 46, No 6Journal of the American Association for Laboratory Animal ScienceNovember 2007
biomarker used to detect promoter activity. However, histologic
15. Dvoroznakova E, Boroskova Z, Dubinsky P, Velebny S, Tomaso-
changes compatible with those of promoter activity were seen
vicova O, Machnicka B. 1998. Changes in cellular immunity of
only at doses exceeding the therapeutic level. In addition, in vivo
mice treated for larval toxocarosis with fenbendazole. Helmint-
and in vitro evidence from rodents and other animal species
hologia 35:189–195.
16. Franke DDH, Shirwan H. 2006. Prophylactic fenbendazole therapy
indicates that, compared with prototype inducers, FBZ moder-
does not affect the incidence and onset of type 1 diabetes in non-
ately induces cytochromes 1A1, 1A2, and 2B1. These isoforms
obese diabetic mice. Int Immunol 18:453–458.
(particularly 1A1) play an important role in activating endog-
17. Gary AT, Kerl ME, Wiedmeyer CE, Turnquist SE, Cohn LA. 2004.
enous (for example, estrogens) and exogenous procarcinogens.
Bone marrow hypoplasia associated with fenbendazole adminis-
The effects of potential pharmacokinetic interactions of other
tration in a dog. J Am Anim Hosp Assoc 40:224–229.
drugs with FBZ, including acceleration of its own CYP2B1-
18. Gleizes-Escala C, Lesca P, Larrieu G, Dupuy J, Pineau T, Galtier P.
mediated oxidation to hydroxyl FBZ, have not been studied.
1996. Effect of exposure of rabbit hepatocytes to sulfur-containing
At present, there are confl icting reports on whether FBZ exerts
anthelmintics (oxfenbendazole and fenbendazole) on cytochrome P4501A1 expression. Toxicol In Vitro 10:129–139.
immunomodulatory actions, and although most studies have
19. Goldenthal EI. 1980. Three-generation reproduction study in
not shown any effects on selected immune responses, whether
rats. Mattawan (MI): International Research and Development
lymphocyte proliferation in vitro is suppressed remains con-
troversial and deserves further consideration. At therapeutic 20. Godenthal EI. 1980. 24-month oral carcinogenicity study in mice. levels, neither reproductive, teratologic, nor behavioral studies
Mattawan (MI): International Research and Development Corpo-
have shown any signifi cant biologic effects of FBZ.
21. Godenthal EI. 1980. Lifetime oral toxicity study in rats. Mattawan
(MI): International Research and Development Corporation. References
22. Gozalo AS, Schwiebert RS, Lawson GW. 2006. Mortality associ-
1. Agersborg SS, Garza KM, Tung KSK. 2001. Intestinal parasit-
ated with fenbendazole administration in pigeons (Columba livia).
ism terminates self tolerance and enhances neonatal induction of
J Am Assoc Lab Anim Sci 45:63–66.
autoimmune disease and memory. Eur J Immunol 31:851–859.
23. Hill WA, Randolph MM, Lokey SJ, Hayes E, Boyd KL, Mandrell Baliharova V, Skalova L, Mass RFM, De Vrieze G, Bull S, Fink- TD. 2006. Effi cacy and safety of topical selamectin to eradicate Gremmels J. 2003. The effects of benzimidazole anthelmintics
pinworm (Syphacia spp.) infections in rats (Rattus norvegicus) and
on P4501A in rat hepatocytes and HepG2 cells. Res Vet Med
mice (Mus musculus). J Am Assoc Lab Anim Sci 45:23–26. 75:61–69.
24. Hiroi T, Miyazaki Y, Kobayashi Y, Imaoka S, Funae Y. 1995.
3. Baliharova V, Velik J, Šavlik M, Szotakova B, LAmka J, Tahotna
Induction of hepatic P450s in rat by essential wood and leaf oils.
L, Skalova L. 2004. The effects of fenbendazole, fl ubendazole and
Xenobiotica 25:457–467.
mebendazole on activities of hepatic cytochromes P450 in pig. J
25. Holden HE, Crider PA, Wahrenburg MG. 1980. Mitotic arrest by
Vet Pharmacol Therap 27:85–90.
benzimidazole analogs in human lymphocytes cultures. Environ
4. Barlow SC, Brown MM, Price HV. 2005. Eradication of Syphacia
Mutagen 2:67–73. muris from food-restricted rats without environmental decontami-
26. Howard LL, Papendick R, Stalis IH, Allen JL, Sutheland-Smith
nation. Contemp Top Lab Anim Sci 44:23–25. M, Zuba JR, Ward DL, Rideout BA. 2002. Fenbendazole and
5. Barron S, Baseheart BJ, Segar TM, Deveraux T, Willford JA. 2000.
albendazole toxicity in pigeons and doves. J Avian Med Surg
The behavioral teratogenic potential of fenbendazole: a medication
16:203–210.
for pinworm infestation. Neurotoxicol Teratol 22:871–877.
27. Huerkamp MJ, Benjamin KA, Webb SK, Pullium JK. 2004. Long-
6. Benchaoui HA, and McKellar QA. 1996. Interaction between
term results of dietary fenbendazole to eradicate Syphacia muris
fenbendazole and piperonyl butoxide: pharmacokinetic and
from rat colonies. Contemp Top Lab Anim Sci 43:35–36.
pharmacodynamic implications. J Pharm Pharmacol 48:753–759.
28. Huerkamp MJ, Kimberley AB, Zitzow LA, Pullium JK, Lloyd
7. Bugarski D, Jovcic G, Katic-Radivojevic S, Petakov M, Krstic JA, Thompson WD, Webb SK, Lehner NDM. 2000. Fenbendazole A, Stojanovic N, Milenkovic P. 2006. Hematopoietic changes
treatment without environmental decontamination eradicates
and altered reactivity of IL-17 in Syphacia obvelata-infected mice.
Syphacia muris from all rats in a large, complex research institution.
Parasitol Int 55:91–97.
Contemp Top Lab Anim Sci 39:9–12.
8. Cabaj W, Stankiewicz M, Jonas WE, Moore LG. 1994. Fenbenda-
29. Hunter RL, Dong-Young C, Kincer JF, Cass WA, Bing G, Gash
zole and its effect on the immune system of the sheep. N Z Vet J
DM. 2007. Fenbendazole treatment may infl uence lipopolysac- 42:216–220.
charide effects in rat brain. Comp Med 57:487-492.
9. Campbell WC. 1990. Benzimidazoles: veterinary uses. Parasitol
30. Jackson TA, Hall JE, Boivin GP. 1998. Ivermectin toxicity in mul-
Today 6:130–133.
tiple mouse lines. Lab Anim Pract 31:37–41.
10. Chen J. 2005. Animal models for acquired bone marrow failure
31. Jacoby RO, Lindsey JR. 1997. Health care for research animals is
syndromes. Clin Med Res 3:102–108.
essential and affordable. FASEB J 11:609–614.
11. Coghlan LG, Lee DR, Psencik B, Weiss D. 1993. Practical and
32. Johnston NA, Bieszcak JR, Verhulst S, Disney KE, Montgomery
effective eradication of pinworms (Syphacia muris) in rats by use
KE, Toth LA. 2006. Fenbendazole treatment and litter size in rats.
of fenbendazole. Lab Anim Sci 43:481–486.
J Am Assoc Lab Anim Sci 45:35–39.
12. Dalvi RR, Gawai KR, Dalvi PS. 1991. Lack of in vivo and in
33. Katiyar SK, Gordon VR, McLaughlin GL, Edlind TD. 1994.
vitro effects of fenbendazole on phase I and phase II biotrans-
Antiprotozoal activities of benzimidazoles and correlations with
formation enzymes in rats, mice and chickens. Vet Hum Toxicol
β-tubulin sequence. Antimicrob Agents Chemother 38:2086–2090. 33:548–551.
34. Keen RG, Macinnis MLM, Guilhardi P, Chamberland KA,
13. Dawson PJ, Gutterdge WE, Gull K. 1984. A comparison of the Church RM. 2005. The lack of behavioral effects of fenbendazole:
interaction of anthelmintic benzimidazoles with tubulin isolated
a medication for pinworms infection. Contemp Top Lab Anim Sci
from mammalian tissue and the parasitic nematode Ascadidia
44:17–23.
Galli. Biochem Pharmacol 33:1069–1074.
35. Krutovskikh VA, Mesnil M, Mazzoleni G, Yamasaki H. 1995.
14. Dey A, Jones JE, Nebert DW. 1999. Tissue and cell type-specifi c
Inhibition of rat liver gap junction intercellular communication
expression of cytochrome P450 1A1 and cytochrome P450 1A2
by tumor-promoting agents in vivo. Lab Invest 72:571–577.
mRNA in the mouse localized in situ hybridization. Biochem
36. Kuribayashi M, Asamoto M, Suzuki S, Hokaiwado N,
Pharmacol 58:525–537. Ogawa K, and Shirai T. 2006. Lack of modifi cation of 2-ami- no-3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx) rat hepatocarcinogenesis by caffeine, a CYP1A2 inducer, points to complex counteracting infl uences. Cancer Lett 232:289–299.
Biological effects of fenbendazole in rodents
37. Lake BG, Renwick AB, Cunninghame ME, Price RJ, Surry D, Ev-
58. Sato K. 1988. Glutathione-S-transferases and hepatocarcinogenesis. ans DC. 1998. Comparison of the effects of some CYP3A and other
Jpn J Cancer Res 79:556–572.
enzyme inducers on replicative DNA synthesis and cytochrome
59. Šavlík M, Fimanova K, Szotakova B, Lamka J, Skalova L. 2006.
P450 isoforms in rat liver. Toxicology 131:9–20.
Modulation of porcine biotransformation enzymes by anthelm-
38. Leung YK, Lau KM, Mobley J, Jiang Z, Ho SM. 2005. Overex-
intic therapy with fenbendazole and fl ubendazole. Res Vet Med
pression of cytochrome P450 1A1 and its novel spliced variant in
80:267–274.
ovarian cancer cells: alternative subcellular enzyme compartmenta-
60. Scholz H and Schultes E. 1973a. Report on an acute oral safety
tion may contribute to carcinogenesis. Cancer Res 65:3726–3734.
evaluation of the anthelmintic HOE 881 in mice. Frankfurt am
39. Löfgren S, Habgjork AL, Ekman S, Fransson-steen R, Terelius Y.
2004. Metabolism of human cytochrome P450 marker substrates in
61. Scholz H and Schultes E. 1973b. Report on an acute oral safety
mouse: a strain and gender comparison. Xenobiotica 34:811–834.
evaluation of the anthelmintic HOE 881 in rats. Frankfurt am Main
40. Mandal PK. 2005. Dioxin: a review of its environmental effects
and its aryl hydrocarbon receptor biology. J Comp Physiol B
62. Shimada T, Oda Y, Gillan EM, Guengerich P, Inoue K. 2001. 175:221–230.
Metabolic activation of polycyclic aromatic hydrocarbons and
41. McNair DM, Timmons EH. 1977. Effects of Aspiculuris tetraptera
other procarcinogens by cytochromes P4501A1 and P4501B1 al-
and Syphacia obvelata on exploratory behavior of an inbred mouse
lelic variants and other human cytochromes P450 in Salmonella
strain. Lab Anim Sci 27:38–42. typhimurium NM2009. Drug Metab Disp 29:1176–1182.
42. Michels C, Goyal P, Nieuwenhuizen N, Brombacher F. 2006.
63. Shirai T. 1997. A medium-term rat liver bioassay as a rapid in
Infection with Syphacia obvelata (pinworm) induces protective Th2
vivo test for carcinogenic potential: a historical review of model
immune responses and infl uences ovoalbumin-induced allergic
development and summary of results from 291 tests. Toxicol Pathol
reactions. Infect Immun 74:5926–5932. 25:453–460.
43. Mitsumori K, Onodera H, Shoda T, Uneyama C, Imazawa T,
64. Shi-Xin X, Ding Z, Yu-Mei S, Shu-Huai W, Li-Qing S. 1992. Sub- Takegawa K, Yasuhara K, Watanabe T, and Takahashi M. 1997.
chronic toxicity studies of fenbendazole in rats. Vet Hum Toxicol
Liver tumor-promoting effects of oxfenbendazole in rats. Food
34:411–413.
Chem Toxicol 35:799–806.
65. Shoda T, Onodera H, Takeda M, Uneyama C, Imazawa T, Takeg-
44. Mohn G, Philipp EM. 1981. Effects of Syphacia muris and the awa K, Yasuhara K, Watanabe T, Hirose M, Mitsumori K. 1999.
anthelmintic fenbendazole on the microsomal monooxygenase
Tumor promoting effects of fenbendazole in rats. Toxicol Pathol
system in mouse liver. Lab Anim 15:89–95. 27:553–562.
45. Moon YJ, Wang X, Morris ME. 2006. Dietary fl avonoids: ef-
66. Short CR, Barker SA, Hsieh LC, Ou SP, McDowell T. 1988. Dis-
fects of xenobiotic and carcinogen metabolism. Toxicol In Vitro
position of fenbendazole in the rabbit. Res Vet Sci 44:215–219. 20:187–210.
67. Short CR, Flory W, Hsieh LC, Barker SA. 1988. The oxidative
46. Murray M, Hudson AM, Yassa V. 1992. Hepatic microsomal
metabolism of fenbendazole: a comparative study. J Vet Pharmacol
metabolism of the anthelmintic benzimidazole fenbendazole:
Therap 11:50–55.
enhanced inhibition of cytochrome P450 reactions by oxidized
68. Silveira AC, Gilioli R, Oliveira ES, Bassani RA. 2002. Subsensitiv-
metabolites of the drug. Chem Res Toxicol 5:60–66.
ity to beta-adrenergic stimulation in atria from rats infested with
47. Nakamato T, Oda Y, Imaoka S, Funae Y, Fujimori M. 1997. Effect Syphacia sp. Lab Anim 37:63–67.
of phenobarbital on the pharmacokinetics of lidocaine monoeth-
69. Stankiewicz M, Cabaj W, Jonas WE, Moore LG, Chie WNG. 1994.
ylglycinexylidide and 3-hydroxylidocaine in the rat: correlation
Oxfenbendazole treatment of non-parasitized lambs and its effect
with P450 isoform levels. Drug Metab Disp 25:296–300.
on the immune system. Vet Res Commun 18:7–18.
48. Neiffer DL, Lydick D, Burks K, Doherty D. 2005. Hematologic
70. Suzuki S, Takahashi S, Asamoto K, Inaguma S, Ogiso T,
and plasma biochemical changes associated with fenbendazole
Hirose M, Shirai T. 2002. Lack of modification of 2-amino-
administration in Hermann’s tortoises (Testudo hermanni). J Zoo
3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx)-induced
Wildlife Med 36:661–672.
hepatocarcinogenesis in rats by fenbendazole – a CYP1A2 inducer.
49. Parish SJ, McFarlane RG, Familton AS, Abell TJ. 1996. The ef-
Cancer Lett 185:39–45.
fect of fenbendazole on the immune system of lambs. Proc New
71. Taffs LF. 1976. Pinworm infections in laboratory rodents: a review.
Zealand Soc Anim Prod56:80–83.
Lab Anim 10: 1–13.
50. Philip M, Rowley DA, Schreiber H. 2004. Infl ammation as a tumor
72. Toth LA, Oberbeck C, Straign CM, Frazier S, Rehg JE. 2000. Toxic-
promoter in cancer induction. Semin Cancer Biol 14:433–439.
ity evaluation of phophylactic treatments for mites and pinworms
51. Pitot HC, Dragan Y, Sargent L, Xu YH. 1991. Biochemical mark-
in mice. Contemp Topics Lab Anim Sci 39:18–21.
ers associated with the stages of promotion and progression
73. Trosko JE, Ruch RJ. 2002. Gap junctions as targets for cancer
during hepatocarcinogenesis in the rat. Environ Health Perspect
chemoprevention and chemotherapy. Curr Drug Targets 3:1–17 93:181–189.
74. Turesky RJ, Constable A, Richoz J, Varga N, Markovic J, Martin
52. Plumb DC. 1999. Veterinary drug handbook, 3rd ed. Ames (IA): MV, Guengerich FP. 1998. Activation of heterocyclic aromatic
amines by rat and human liver microsomes and by purifi ed rat
53. Pritchett KR, Johnston NA. 2002. A review of treatments for the
and human cytochrome P4501A2. Chem Res Toxicol 11:925–936.
eradication of pinworm infections from laboratory rodent colonies.
75. Wagner M. 1988. The effect of infection with the pinworm (Syphacia
Contemp Top Lab Anim Sci 41: 36–46. muris) on rat growth. Lab Anim Sci 38:476–478.
54. Redbook 2000 [Internet]. Toxicological principles for the safety
76. Weber MA, Miller MA, Neiffer DL, Terrell SP. 2006. Presumptive
assessment of food ingredients: 2007 update [cited 26 Sep 2007].
fenbendazole toxicosis in North Amercian porcupines. J Am Vet
Available at http://www.cfsan.fda.gov/~redbook/red-ivc6.
Med Assoc 228:1240–1242.
77. World Health Organization [Internet]. WHO food additive series,
55. Reiss CS, Herrman JM, Hopkins RE. 1987. Effect of anthelm-
no. 29. Toxicological evaluation of certain veterinary drug residues:
inthic treatment on the immune response of mice. Lab Anim Sci
1991 [cited 26 Sep 2007]. Available at http://www.inchem.org/
37:773–775.
documents/jecfa/jecmono/v29je01.htm.
56. Sajid MS, Muhammad ZIG, Iqbal MU. 2006. Immunomodu-
78. Zaias, J. 2007. Personnel communication.
latory effect of various anti-parasitics: a review. Parasitology
79. Zhao LZ, Chen J, Ee PLR, Chan E, Duan W, Guan YY, Hong 132:301–313. YH, Chen X, Zhou S. 2006. Induction of propanolol metabo-
57. Samson-Himmelstjerna G, Witzendorff C, Sievers G, Schnieder
lism by ginkgo biloba extract EGb 761 in rats. Curr Drug Metab
T. 2002. Comparative use of faecal egg count reduction test, egg 7:577–587.
hatch assay, and beta-tubulin codon 200 genotyping in small strongyles (cyathostominae) before and after benzimidazole treat- ment. Vet Parasitol 108:227–235.
J CATARACT REFRACT SURG - VOL 32, JANUARY 2006Transient light sensitivity after femtosecond laserflap creation: Clinical findings and managementKarl G. Stonecipher, MD, Jon G. Dishler, MD, Teresa S. Ignacio, MD, Perry S. Binder, MDPURPOSE: To describe the constellation of subjective and objective findings associated with unusualoccurrences of photosensitivity after laser in situ keratomileusis
Ist die vorsorglicher Entwurmung von Pferden sinnvoll? Ist die vorsorgliche Entwurmung von Pferden sinnvoll? von Sabine Müller In vielen Reitställen ist eine vorsorgliche chemische Entwurmung aller Pferde drei bis vier Mal im Jahr üblich. Dabei wird vorher nicht untersucht, ob und in welchem Ausmaß ein Wurmbefall vorliegt und es wird auch nicht kontrolliert, ob die Entwurmung erfolgreich war.
 Vol 46, No 6Journal of the American Association for Laboratory Animal ScienceNovember 2007
connexin 32 is a necessary but insuffi cient factor to consider a compound as a tumor promoter. Two things are necessary for an initiated cell to proliferate: a) an intracellular signal from the chemical to block contact inhibition and transfer of signals from cell to cell via gap junctions, and b) an intracellular signal to proliferate. Disruption of gap junctions does not necessarily imply that the chemical induces a mitogenic signal.
A previous study looked at another potential biomarker of tu-
mor promotion: the induction of certain cytochromes P450 such as CYP2B1.65 These are discussed later in the text. Nevertheless, that a compound exerts an inducing effect on cytochrome P450s may not necessarily imply a promoter or mitogenic action; and so, induction should not be considered a reliable endpoint of promoter activity.37 This point is illustrated by the induction of CYP1A2 by FBZ in relation to a known dietary carcinogen, MeIQx (2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline). MeIQx is an heterocyclic amine thought to be metabolically bioactivated to genotoxic intermediates in the liver by CYP1A2.36,74 Combining FBZ (600 ppm in the diet) with MeIQx did not enhance MeIQx-induced hepatocarcinogenesis, despite the fact that FBZ caused a 2.3-fold increase in CYP1A2 levels.70 In this case, only GST-P positive liver cell foci were used to assess carcinogenicity.
When the same 2-stage carcinogenesis model was applied
to OXF, which is the primary in vivo metabolite of FBZ, lower doses of OXF (10 and 100 ppm) had greater effects at inducing the same cytochromes and affecting the same assays (connexin 32 and GST-P) used as biomarkers of preneoplastic lesions.43 The higher potency of OXF suggests that many of the in vivo effects of FBZ likely were caused through this metabolite.
Vol 46, No 6Journal of the American Association for Laboratory Animal ScienceNovember 2007
connexin 32 is a necessary but insuffi cient factor to consider a compound as a tumor promoter. Two things are necessary for an initiated cell to proliferate: a) an intracellular signal from the chemical to block contact inhibition and transfer of signals from cell to cell via gap junctions, and b) an intracellular signal to proliferate. Disruption of gap junctions does not necessarily imply that the chemical induces a mitogenic signal.
A previous study looked at another potential biomarker of tu-
mor promotion: the induction of certain cytochromes P450 such as CYP2B1.65 These are discussed later in the text. Nevertheless, that a compound exerts an inducing effect on cytochrome P450s may not necessarily imply a promoter or mitogenic action; and so, induction should not be considered a reliable endpoint of promoter activity.37 This point is illustrated by the induction of CYP1A2 by FBZ in relation to a known dietary carcinogen, MeIQx (2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline). MeIQx is an heterocyclic amine thought to be metabolically bioactivated to genotoxic intermediates in the liver by CYP1A2.36,74 Combining FBZ (600 ppm in the diet) with MeIQx did not enhance MeIQx-induced hepatocarcinogenesis, despite the fact that FBZ caused a 2.3-fold increase in CYP1A2 levels.70 In this case, only GST-P positive liver cell foci were used to assess carcinogenicity.
When the same 2-stage carcinogenesis model was applied
to OXF, which is the primary in vivo metabolite of FBZ, lower doses of OXF (10 and 100 ppm) had greater effects at inducing the same cytochromes and affecting the same assays (connexin 32 and GST-P) used as biomarkers of preneoplastic lesions.43 The higher potency of OXF suggests that many of the in vivo effects of FBZ likely were caused through this metabolite.