La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Pico.sssup.it
J. Plant Physiol. 159. 1383 – 1386 (2002)
Urban & Fischer Verlaghttp://www.urbanfischer.de/journals/jpp
Short Communication Elicitors of defence responses repress a gibberellin signalling pathway in barley embryos
Elena Loreti1, Daniela Bellincampi2, Christel Millet3, Amedeo Alpi3, Pierdomenico Perata4 *
1 Istituto di Biologia e Biotecnologia Agraria, sezione di Pisa, Via del Borghetto 80, 56100 Pisa, CNR, Italy2 Dipartimento di Biologia Vegetale Universita’ di Roma «La Sapienza», P. le A. Moro 5, 00185 Roma, Italy3 Department of Crop Plant Biology, University of Pisa, Via Mariscoglio 34, 56124 Pisa, Italy4 Department of Agricultural Sciences, University of Modena and Reggio Emilia, Via Kennedy 17, 42100 Reggio Emilia, Italy
Received January 31, 2002 · Accepted April 22, 2002
Elicitors of defence response can modulate pathways other than those related to pathogen attacks. In this paper, we demonstrate that, as with sugars, chitosan and oligogalacturonides (OGs) can re-press the gibberellin signalling pathway leading to the induction of α-amylase in barley embryos. These results are suggestive of a complex cross-talk between the defence, hormonal and metabolicsignalling pathways. Key words: α-amylase – barley – chitosan – oligogalacturonides – sugar repression Abbreviations: OGs = Oligogalacturonides. – GA =
Gibberellic acid. – DP = Degree of polimeriza-
Introduction
bolic regulation of gene expression, implying that the actionof hormones requires an adequate metabolic status within the
Plants are subjected to a wide variety of internal and external
plant cell, reflected by its sugar content. This is not surprising
stimuli, including the classical hormonal modulation of plant
since the cross-talk between hormonal and other signalling
development and adaptation to the environment, such as the
pathways have been demonstrated (Gazzarini and McCourt
response to pathogen attack. Intriguingly, these signalling
pathways do not appear to act independently of each other,
Environmental signals such as wounding and pathogen
and connections between signalling pathways are feasible.
attack can alter the response to hormones; elicitors of plant
An example of interaction between signalling pathways is
defence response such as oligogalacturonides (Ridley et al.
revealed by the cross-talk between the hormonal and meta-
2001) can catalyse the response to auxin in tobacco leafexplants (Bellincampi et al. 1993, 1996). Intriguingly, sugars
* E-mail corresponding author: perata.pierdomenico@unimo.it
can act as signalling molecules triggering the induction of
defence-related genes (Herbers and Sonnewald 1998) and
tal RNA loaded in electrophoresis was 20 µg. RNA was electropho-
elicitors of defence responses can modulate the expression
resed and blotted on nylon membrane (BrightStar-Plus, Ambion) by
of sugar-regulated genes (Ehness et al. 1997). It is thus tempt-
using the procedure suggested by the manufacturer. Membranes
were pre-hybridised and hybridised using the NorthernMax kit (Am-
ing to speculate that the hormonal, metabolic, and defence
bion). Radiolabelled probes were prepared from gel-purified cDNA in-
signalling pathways share common elements along their
serts by random primer labelling (Takara Chemicals) with [α32P]-
respective signalling pathways. In this study, we investigated
dCTP. Equal loading was checked by re-probing with a barley ubiqui-
the effects of elicitors of plant defence response on the hor-
tin cDNA probe. Statistical significance of the data reported in the
monal induction of α-amylase in cereal grains, which is one of
RNA gel blots was checked by analysing at least three replicate ex-
the model systems with which to study hormone action
periments and their quantitative, ubiquitin-normalised data after
(Bethke et al. 1997). It has been demonstrated that sugars
can act as signals whose presence interferes with the gibbe-rellin-signalling leading to sugar-repression of the induction ofα-amylase (Perata et al. 1997). In Chenopodium rubrum sus-
Results and Discussion
pension cultures, the fungal elicitor, chitosan, is able to trig-ger gene modulation mirroring that elicited by glucose
Glucose and the fungal elicitor chitosan have been shown to
(Ehness et al. 1997). Assuming that common elements exist in
display a co-ordinated regulatory mechanism. In Chenopo-
the hormonal, metabolic, and defence signalling pathways,
dium rubrum both signals induce the expression of the defen-
elicitors of plant defence response should be able to mimic
ce-related phenylalanine ammonia-lyase (PAL), the sink-spe-
the effects of sugars on the hormonally modulated α-amylase
cific extracellular invertase (CIN1) genes and the repression
gene. In this paper this hypothesis is investigated.
of ribulose bisphosphate carbossilase (RbcS) gene (Ehnesset al. 1997).
Gibberellin-treated barley embryos produce α-amylase
Materials and Methods
mRNA and glucose strongly interferes with this process (Pe-rata et al. 1997). We analysed the effect of two plant defence
Plant material
Barley (Hordeum vulgare L.) grains of cv. Himalaya were used. Em-
bryos were dissected from sterilised grains (shaken in 5 % sodium hy-
pochlorite for 1 h; washed in sterile water with shaking for 2 h) using a
scalpel as previously described (Perata et al. 1997). Incubation of em-
bryos was carried out at 25 ˚C with vigorous shaking in 24-well plastic
plates, each well containing four embryos and 500 µL of 5 mmol/LCaCl2 with 5 µg of chloramphenicol. When used, GA3 (1µmol/L), chito-san, or oligogalacturonides were added. Chemicals
The commercially available compounds were purchased from Sigma.
OGs with a DP from 1 to 18 were generated by partial digestion of
polygalacturonic acid (Na + salt) with homogeneous Aspergillus nigerendopolygalacturonase, and the mixture of different sized OGs was
separated to obtain two preparations of OGs with degree of polimeri-
sation (DP) of 1 to 8 and 9 to 18, respectively, as previously described
cDNA probes
The high-pI α-amylase probe was clone pM/C (Rogers 1985); theprobe for ubiquitin was a barley probe detecting different size mes-
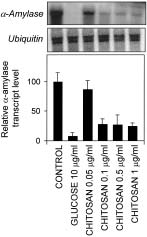
Figure 1. Glucose and chitosan repress α-amylase in barley embryos.
sengers of the ubiquitin multigene family (Gausing and Barkardottir
Embryos were incubated in 5 mmol/L CaCl2, and 1µmol/L GA3, 10 µg/
mL glucose or chitosan (0.05 –1 µg/mL) for 24 h. RNA was extractedfrom embryos for gel blot analysis. Blots were probed with the α-amy-lase probe and reprobed with an ubiquitin probe. RNA levels of bothα
RNA isolation and gel blots
-amylase and ubiquitin were quantified and the «Relative α-amylase
transcript level» normalised to ubiquitin (100 = transcript level in the
RNA extraction was performed by using the aurintricarboxylic acid
embryos not treated with exogenous glucose or chitosan) is reported
method as previously described (Perata et al. 1997). The amount of to-
in the histograms. Data are mean ± SE of 3 separate experiments.
Repression of α-amylase by defence elicitors
elicitors such as chitosan and oligogalacturonides (OGs) on
tional activation of defence-related genes such as those en-
the induction of α-amylase mRNA by gibberellic acid. Figure
coding for phenylalanine ammonia-lyase, chalcone synthase,
1 shows that chitosan is able to repress the inductive process
and the pathogenesis-related proteins PR-1 and PR-2 (De Lo-
leading to α-amylase mRNA accumulation. As a control, rep-
renzo et al. 1987, Messiaen and Van Cutsem 1993). OGs have
ression by glucose (10 µg/mL) is also shown. The chitosan
been reported to affect several aspects of plant growth and
concentrations (0.05 –1 µg/mL) able to trigger repression of
development as well (Altamura et al. 1998, Branca et al. 1988,
α-amylase mRNA are in the same range as reported by Eh-
Bellincampi et al. 1993, 1996). The majority of defence re-
ness et al. (1997) in Chenopodium rubrum cells.
sponses and developmental effects exerted by oligogalactu-
α-1,4-linked oligogalacturonides (OGs) derived from the
ronides requires a degree of polymerispolymerisation (DP)
plant cell wall have been shown to elicit several defence re-
between 10 and 15, even though smaller oligomers with a DP
sponses (Côté and Hahn 1994) and to induce the transcrip-
less than 8 have been shown to activate defence responsesin plants also (Côté and Hahn 1994). OGs with a degree ofpolymerisation of 9 –18 (DP 9 –18) used at concentrationhigher than 1µg/mL trigger repression of α-amylase mRNA in-duced by gibberellin in barley embryos (Fig. 2 A, C). In con-trast incubation of embryos with short OGs (DP 1– 8) (Fig. 2 B, C) resulted in no inhibitory effect of α-amylase ex-pression. The effect of OGs on α-amylase belongs to thelong-size OG-dependent responses. Although the reason forthe observed long-size dependence of the same responsesto OGs is not known, indirect evidence indicates that the for-mation of intermolecular dimeric or multimeric polygalacturo-nate chains cross-linked to Ca2+ ions are the active molecularsignals (Messiaen and Van Cutsem al. 1994).
Plant responses induced by OGs usually have been inves-
tigated in dicot plants. It is noteworthy that we have shown aresponse to OGs in barley, indicating the presence of a per-ception and transduction mechanism sensitive to these oligo-saccharines in monocots. In many systems utilised in previ-ous analyses, including suspension-cultured cells, proto-plasts or leaf explants, OGs were likely to have either facili-tated or direct access to the cells or damaged tissues. In thiswork, we utilised intact barley embryos, showing that OGs arereadily taken up by plants. Moreover, the use of intact em-bryos prevents the possibility of rapid fragmentation of theOGs added to the incubation medium due to extracellularpolygalacturonase released by cultured cells or by woundedtissue (Mathieu et al. 1998, Bergey et al. 1999). Chitosan(1 µg/mL), OGs with DP 9–18 (8 µg/mL) did not affect germi-nation of the isolated barley embryos (not shown) ruling outthe possibility of lower α-amylase expression as a side effectof toxicity in these elicitors on the germination process.
α-Amylase has been shown to be a late gibberellin-
induced gene, requiring about 12 –15 h for its expression to beevident (Fig. 3 A). When chitosan and OGs with DP 9 –18 were
Figure 2. Oligogalacturonides repress α-amylase in barley embryos.
added 9 h after treatment with gibberellins (corresponding to
A, embryos were incubated in 5 mmol/L CaCl2, 1 µmol/L GA3, and
time 0 in Fig. 3 B) and just before the sharp rise in α-amylase
increasing concentrations of OGs with DP 9 –18 (0 – 8 µg/mL) for 24 h;
transcript, the resultant inhibition of gene expression was of
B, embryos were incubated in 5 mmol/L CaCl2, 1 µmol/L GA3, and
the same magnitude as that observed by adding the plant
increasing concentrations of OGs with DP 1– 8 (0 –10 µg/mL) for 24 h.
elicitors from the beginning (see Fig. 2). This indicates that
RNA was extracted from embryos for gel blot analysis. Blots were
the effect on gene expression is quite rapid. Similar con-
probed with the α-amylase probe and reprobed with an ubiquitinprobe.
clusions have been reported previously by Bellincampi et al. C, RNA levels of both α-amylase and ubiquitin were quantified
and the «Relative α-amylase transcript level» normalised to ubiquitin
(2000), who treated tobacco leaf explants with the same OG
(100 = transcript level in the embryos not treated with OGs) is
mixtures. These authors showed, in fact, that OGs could rep-
reported in the line graph. Data are from a representative experiment.
ress the induction of the late auxin-induced rolB gene even
viability of embryos was not compromised by accumulation ofreactive oxygen species. This is also supported by the lack oftoxic effects of fungal elicitors on the germination of isolatedbarley embryos (data not shown).
As discussed by Herbers et al. (1996), pathogen infection
may affect the sugar status of the infected plant tissue,usually with elevated sugar levels. Remarkably, Herbers et al. (1996) demonstrated that up-regulation of extracellular inver-tase, leading to higher hexose levels in the apoplast, resultsin defence responses and systemically acquired resistance.
Overall, the results reported here strengthen the hypothe-
sis of the existence of a network of signalling pathways, in-cluding the hormonal, metabolic and defence responses. Thecross-talk within this signalling network results in the induc-tion/repression of genes as the outcome of the perception ofmultiple signals. Acknowledgements. This work was supported by MURST-COFIN Figure 3. Late addition of OGs and chitosan repress α-amylase in barley embryos. A, time course of α-amylase mRNA accumulation fol- lowing addition of GA3. Embryos were incubated in 5 mmol/L CaCl2, References
and 1 µmol/L GA3 for up to 15 h. RNA was extracted from embryos forgel blot analysis. Blots were probed with the α-amylase probe and
Altamura MM, Zaghi D, Salvi G, De Lorenzo G, Bellincampi D (1998)
re-probed with an ubiquitin probe, confirming homogeneous electro-
phoresis loading (not shown). B, embryos were incubated in 5 mmol/L
Bellincampi D, Cardarelli M, Zaghi D, Serino G, Salvi G, Gatz C, Cer-
CaCl2, and 1 µmol/L GA3 for up to 15 h. Various chemicals, as indi-
vone F, Altamura MM, Costantino P, De Lorenzo G (1996) Plant Cell
cated in the figure, were added 9 h after GA3 addition (Time 0). OGs
concentration was 10 µg/mL, chitosan (1 µg/mL), and Actinomycin D
Bellincampi D, Dipierro N, Salvi G, Cervone F, De Lorenzo G (2000)
(ActD, 10 µg/mL). Embryos were extracted for RNA gel blot analysis.
Blots were probed with the α-amylase probe and re-probed with an
Bellincampi D, Salvi G, De Lorenzo G, Cervone F, Marfà V, Eberhard
ubiquitin probe, confirming homogeneous electrophoresis loading
S, Darvill A, Albersheim P (1993) Plant J 4: 207– 213
Bergey DR, Orozco-Cardenas M, De Moura DS, Ryan CA (1999) Proc
Bethke P, Schuurink R, Jones RL (1997) J Exp Bot 48: 1337–1356
when applied several hours after auxin treatment (Bellincampi
Branca C, De Lorenzo G, Cervone F (1988) Physiol Plant 72: 499 – 504
et al. 2000). OGs with DP 1– 8, added 9 hours after gibberellin
Côté F, Hahn MG (1994) Plant Mol Biol 26: 1379 –1411
treatment are unable to prevent the increase in α-amylase
De Lorenzo G, Ranucci A, Bellincampi D, Salvi G, Cervone F (1987)
expression (Fig. 3 B), indicating their inactivity was not due to
Ehness R, Ecker M, Godt DE, Roitsch TH (1997) Plant Cell 9: 1825 –
their fragmentation in the culture medium. The effect of chito-
san and OGs with DP 9 –18 suggests a transcriptional block
Fath A, Bethke PC, Jones RL (2001) Plant Physiol 156 –166
mimicking the effect of the transcriptional inhibitor actinomy-
Fath A, Bethke PC, Lonsdale J, Meza-Romero R, Jones RL (2000)
Oligogalacturonides induce accumulation of extracellular
Gausing K, Barkardottir R (1986) Eur J Biochem 158: 57– 62
H2O2, though this is not involved in the inhibition of the auxin-
Gazzarini S, McCourt P (2001) Curr Opin Plant Biol 4: 387– 391
regulated rolB gene expression in tobacco leaf explants
Herbers K, Meuwly P, Frommer WB, Métreaux JP, Sonnewald U (1996)
(Bellincampi et al. 2000). Interestingly, reactive oxygen spe-
cies are responsible for cell death in barley aleurones, a tis-
Herbers K, Sonnewald U (1998) J Plant Res 111: 323 – 328
Mathieu Y, Guern J, Spiro MD, O’Neill MA, Kates K, Darvill AG, Albers-
sue able to respond to GA3 in terms of α-amylase production
(Fath et al. 2001). Cell death occurs after the induction ofα
Messiaen J, Van Cutsem P (1993) Plant Cell Physiol 34: 1117–1123
-amylase has taken place in barley aleurones (Fath et al.
Messiaen J, Van Cutsem P (1994) Plant Cell Physiol 35: 677– 689
2000), indicating that H2O2 endogenously produced is un-
Perata P, Matsukura C, Vernieri P, Yamaguchi J (1997) Plant Cell 9:
likely to affect induction of starch degradation enzymes. Oli-
gogalacturonides-induced H2O2 production could not be de-
Ridley BL, O’Neill MA, Mohnen D (2001) Phytochemistry 57: 929 – 967
tected in barley embryo (data not shown), suggesting that
Rogers JC (1985) J Biol Chem 260: 3731– 3738
April 28, 2008 Rise of Nationalism DOW JONES REPRINTS Frays Global Ties non-commercial use only. To order presentation-ready copies for Trade, Environment distribution to your colleagues, clients or customers, use the Order Face New Threats; Balkanized Internet By BOB DAVIS April 28, 2008; Page A1 format. • Order a reprint of this article now. The w
The Blood and Marrow Transplant Program atOhio State University Comprehensive Cancer Center -James Cancer Hospital and Solove Research Institutehas recently been named as a Blue DistinctionCenter for Transplantation of Adult Autologous andAllogeneic hematopoietic cells. Previously we had Steven M. Devine MD only been awarded this distinction for recipients of Director, Blood and Marrow
 defence-related genes (Herbers and Sonnewald 1998) and
tal RNA loaded in electrophoresis was 20 µg. RNA was electropho-
elicitors of defence responses can modulate the expression
resed and blotted on nylon membrane (BrightStar-Plus, Ambion) by
of sugar-regulated genes (Ehness et al. 1997). It is thus tempt-
using the procedure suggested by the manufacturer. Membranes
were pre-hybridised and hybridised using the NorthernMax kit (Am-
ing to speculate that the hormonal, metabolic, and defence
bion). Radiolabelled probes were prepared from gel-purified cDNA in-
signalling pathways share common elements along their
serts by random primer labelling (Takara Chemicals) with [α32P]-
respective signalling pathways. In this study, we investigated
dCTP. Equal loading was checked by re-probing with a barley ubiqui-
the effects of elicitors of plant defence response on the hor-
tin cDNA probe. Statistical significance of the data reported in the
monal induction of α-amylase in cereal grains, which is one of
RNA gel blots was checked by analysing at least three replicate ex-
the model systems with which to study hormone action
periments and their quantitative, ubiquitin-normalised data after
(Bethke et al. 1997). It has been demonstrated that sugars
can act as signals whose presence interferes with the gibbe-rellin-signalling leading to sugar-repression of the induction ofα-amylase (Perata et al. 1997). In Chenopodium rubrum sus-
Results and Discussion
defence-related genes (Herbers and Sonnewald 1998) and
tal RNA loaded in electrophoresis was 20 µg. RNA was electropho-
elicitors of defence responses can modulate the expression
resed and blotted on nylon membrane (BrightStar-Plus, Ambion) by
of sugar-regulated genes (Ehness et al. 1997). It is thus tempt-
using the procedure suggested by the manufacturer. Membranes
were pre-hybridised and hybridised using the NorthernMax kit (Am-
ing to speculate that the hormonal, metabolic, and defence
bion). Radiolabelled probes were prepared from gel-purified cDNA in-
signalling pathways share common elements along their
serts by random primer labelling (Takara Chemicals) with [α32P]-
respective signalling pathways. In this study, we investigated
dCTP. Equal loading was checked by re-probing with a barley ubiqui-
the effects of elicitors of plant defence response on the hor-
tin cDNA probe. Statistical significance of the data reported in the
monal induction of α-amylase in cereal grains, which is one of
RNA gel blots was checked by analysing at least three replicate ex-
the model systems with which to study hormone action
periments and their quantitative, ubiquitin-normalised data after
(Bethke et al. 1997). It has been demonstrated that sugars
can act as signals whose presence interferes with the gibbe-rellin-signalling leading to sugar-repression of the induction ofα-amylase (Perata et al. 1997). In Chenopodium rubrum sus-
Results and Discussion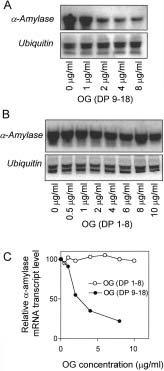 Repression of α-amylase by defence elicitors
elicitors such as chitosan and oligogalacturonides (OGs) on
tional activation of defence-related genes such as those en-
the induction of α-amylase mRNA by gibberellic acid. Figure
coding for phenylalanine ammonia-lyase, chalcone synthase,
1 shows that chitosan is able to repress the inductive process
and the pathogenesis-related proteins PR-1 and PR-2 (De Lo-
leading to α-amylase mRNA accumulation. As a control, rep-
renzo et al. 1987, Messiaen and Van Cutsem 1993). OGs have
ression by glucose (10 µg/mL) is also shown. The chitosan
been reported to affect several aspects of plant growth and
concentrations (0.05 –1 µg/mL) able to trigger repression of
development as well (Altamura et al. 1998, Branca et al. 1988,
α-amylase mRNA are in the same range as reported by Eh-
Bellincampi et al. 1993, 1996). The majority of defence re-
ness et al. (1997) in Chenopodium rubrum cells.
Repression of α-amylase by defence elicitors
elicitors such as chitosan and oligogalacturonides (OGs) on
tional activation of defence-related genes such as those en-
the induction of α-amylase mRNA by gibberellic acid. Figure
coding for phenylalanine ammonia-lyase, chalcone synthase,
1 shows that chitosan is able to repress the inductive process
and the pathogenesis-related proteins PR-1 and PR-2 (De Lo-
leading to α-amylase mRNA accumulation. As a control, rep-
renzo et al. 1987, Messiaen and Van Cutsem 1993). OGs have
ression by glucose (10 µg/mL) is also shown. The chitosan
been reported to affect several aspects of plant growth and
concentrations (0.05 –1 µg/mL) able to trigger repression of
development as well (Altamura et al. 1998, Branca et al. 1988,
α-amylase mRNA are in the same range as reported by Eh-
Bellincampi et al. 1993, 1996). The majority of defence re-
ness et al. (1997) in Chenopodium rubrum cells.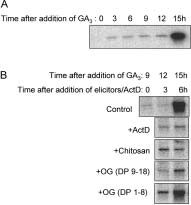 viability of embryos was not compromised by accumulation ofreactive oxygen species. This is also supported by the lack oftoxic effects of fungal elicitors on the germination of isolatedbarley embryos (data not shown).
viability of embryos was not compromised by accumulation ofreactive oxygen species. This is also supported by the lack oftoxic effects of fungal elicitors on the germination of isolatedbarley embryos (data not shown).