La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
9k049901506p
Subtype-Specific Effects of Lithium on Glutamate Receptor Function
NIKOLAS B. KARKANIAS1 AND ROGER L. PAPKE1,21Department of Neuroscience and 2Department of Pharmacology and Therapeutics, University of Florida Medical College,J. H. Miller Health Center, Gainesville, Florida 32610-0267Karkanias, Nikolas B. and Roger L. Papke. Subtype-specific ef-
how synaptic activity and neuronal plasticity may be coupled
fects of lithium on glutamate receptor function. J. Neurophysiol. 81:
1506 –1512, 1999. We report that substitution of sodium with lithium
Several subtypes of GluRs contribute to fast excitatory trans-
(Liϩ) in the extracellular solution causes subtype-specific changes in
mission, and they can be pharmacologically distinguished into
the inward and outward currents of glutamate receptors (GluRs),without a shift in reversal potential. Liϩ produces an increase of
two major classes, non–N-methyl-D-aspartate (NMDA) and
inward and outward currents of ␣-amino-3-hydroxy-5-methyl-4-isox-
NMDA sensitive. The non-NMDA–sensitive channels contain
azole propionate receptors and decreases in the currents of kainate
the receptor subunits GluR1–GluR7. The channels composed
(KA) and N-methyl-D-aspartate receptors. The greatest effect of Liϩ
of subunits GluR5–GluR7 can assemble with accessory sub-
was observed with GluR3. A concentration-response curve for GluR3
units KA1 or KA2 to form receptors that are activated by
reveals that the potentiation caused by Liϩ is greatest at saturating
kainate (KA). Channels composed of GluR1–GluR4 are acti-
agonist concentrations. GluR1, which shows no potentiation by Liϩ at
vated by ␣-amino-3-hydroxy-5-methyl-4-isoxazole propionate
100 M KA, shows a small but significant potentiation at saturatingKA and glutamate concentrations. The effects of Liϩ on outward
(AMPA) as well as KA. The NMDA-sensitive channels are
current, where Liϩ is not the primary charge carrier, and the lack of
composed of NMDAR1 and NMDAR2a–NMDAR2d. Func-
reversal potential shift argue for a mechanism of potentiation not
tional properties of these GluRs, such as their permeability to
associated with Liϩ permeation. This potentiation of current is spe-
sodium, potassium, and calcium and their kinetics, are influ-
cific for Liϩ because rubidium, although causing an increase of
enced by the specific subunit composition of the channel (Holl-
inward current, shifted the reversal potential and did not increase
mann et al. 1991; Monyer et al. 1992) .
outward current. The effects of Liϩ are different for KA, a weak
Receptor desensitization is another property of GluRs that
desensitizing agonist, and glutamate, a strong desensitizing agonist,suggesting that Liϩ might interact with a mechanism of desensitiza-
may regulate synaptic function. When the glutamate transient
tion. By using cyclothiazide (CTZ) to reduce desensitization of
time course is slow, because of the nature of the synaptic
GluR3, we find that for low concentrations of KA and glutamate
morphology, the duration of synaptic current may be deter-
potentiation of the response by a combination of CTZ and Liϩ is no
mined primarily by desensitization kinetics (Barbour et al.
greater than by CTZ or Liϩ alone. However, at high concentrations of
1994). Desensitization is promoted by agonist exposure, and
agonist, the potentiation of the response by a combination of CTZ and
experimental agonists can vary in their relative desensitizing
Liϩ is significantly greater than by CTZ or Liϩ alone. This potenti-
effect. For example, KA produces less desensitization in
ation was additive for glutamate but not for KA. At high agonist
AMPA-selective receptors than either AMPA or glutamate.
concentration in the presence of CTZ, the intrinsically lower desen-
The desensitization kinetics vary among the specific AMPA
sitization produced with KA-evoked responses may preclude Liϩ
receptor subtypes as a result of RNA editing and alternative
from potentiating the current to the same degree as it can potentiateglutamate-evoked responses. The additive effects of CTZ and Liϩ
splicing. The flip/flop domain is a 38-amino acid cassette
were unique to the flop variant of GluR3.
located extracellularly and N-terminal to the final transmem-brane domain of AMPA receptors. Alternative splicing of thiscassette yields mature flip or flop AMPA receptors that vary intheir desensitization kinetics (Mosbacher et al. 1994; Sommer
et al. 1990). Immediately before the flip/flop domain is the R/G
Ionotropic glutamate receptors (GluRs) are responsible for
site (Lomeli et al. 1994). RNA editing at the R/G site can also
most of the fast excitatory neurotransmission in the mamma-
influence AMPA receptor desensitization kinetics with edited
lian brain, which includes activity-dependent synaptic modifi-
channels (G) recovering from desensitization faster.
cations such as long-term potentiation and long-term depres-
Although pharmacological modulators such as cyclothiazide
sion (Bliss and Gardner-Medwin 1973; Bliss and Lynch 1988;
(CTZ) are thought to reduce desensitization of AMPA recep-
Collingridge and Bliss 1987). A differential expression of
tors thus providing pharmacological tools with which to study
GluR subunit genes gives rise to the functional diversity of
this property (Partin et al. 1993; Vyklicky et al. 1991; Wong
GluRs among brain regions with unique permeability and
and Mayer 1993), desensitization was largely assumed to be
kinetic properties for specific receptor subtypes. Characteris-
independent of the charge-carrying ion. In a previous study we
tics of GluRs such as ionic selectivity and kinetics are vital to
reported the preliminary observation that Liϩ produced sub-
the understanding of fast excitatory synaptic transmission and
type-specific alterations of macroscopic current (Karkanias etal. 1998). Further investigation of Liϩ effects on GluR leads usto propose that modulation of receptor desensitization is a
The costs of publication of this article were defrayed in part by the payment
mechanism that causes the flop variant of GluR3 to display a
of page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
modified conductance in the presence of Liϩ.
0022-3077/99 $5.00 Copyright 1999 The American Physiological Society
LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
and then diluted in Ringer to Ͻ1% DMSO. No effect on controlresponse was observed when the agonist was dissolved in DMSO. In
most experiments, barium was used instead of calcium in the Ringer
Female Xenopus laevis frogs were purchased from Nasco (Fort
to minimize contributions of endogenous calcium-activated chloride
Atkinson, WI) and kept in tanks at 17°C. Frogs were anesthetized for
current. However, similar results were obtained in the presence of
30 min on ice in 2.2 g/1.5 l of 3-aminobenzoic acid ethyl ester
calcium. For experiments with GluR6(Q/R), a 2.0-ml, 10-s pulse of
purchased from Sigma (St. Louis, MO). Oocytes were obtained in
concanavalin A (Sigma; St. Louis, MO, type IV; 1.2 mg/ml) was
lobes through a small abdominal incision made just above the leg and
applied 5 min before applying agonist.
near the midline on the ventral surface of the frog. Two to three lobes
Current-voltage relationships were performed by delivering a volt-
were pulled from the frog, cut, and placed in collagenase from
age ramp, Ϫ50.0 mV to ϩ50.0 mV, during the plateau phase of the
Worthington Biochemical (Freehold, NJ) (1 mg/ml in calcium-free
response to agonist (pClamp 5.5, Axon Instruments; Foster City, CA).
Barth’s solution containing 88 mM NaCl, 1 mM KCl, 15 mM HEPES,
The passive current-voltage response of the cell membrane in the
0.33 mM MgSO , and 0.1 mg/ml gentamicin sulfate, pH 7.6) for 2 h
absence of agonist was subtracted from the current-voltage response
to enzymatically remove the native follicular cell layer. After the
in the presence of agonist. Permeability ratios for Liϩ and rubidium
follicular cell layer was removed, oocytes were washed several times
with respect to sodium were calculated with Eq. 1
with calcium-free Barth’s and then washed several times with Barth’s
solution containing 88 mM NaCl, 1 mM KCl, 15 mM HEPES, 0.33
mM CaNO , 0.41 mM CaCl , 0.33 mM MgSO , and 0.1 mg/ml
E is the reversal potential in the presence of X (Liϩ or Rbϩ) or Naϩ,
gentamicin sulfate (pH 7.6) and stored at 17°C. Mature oocytes were
⌬E is the difference between the reversal potentials in X and Na, [X ]
injected the same or following day with the appropriate RNA tran-
is the concentration of X outside of the cell (115 mM), [Na ] is the
concentration of Naϩ outside of the cell (115 mM), and P is thepermeability of the ion.
cDNA clones containing the appropriate gene and T3/T7 bacterial
Concentration-response relationships
promoters were isolated from bacteria and purified with a kit from
The responses of GluR3 expressing oocytes to various test concen-
Qiagen (Santa Clarita, CA). Purified cDNA clones were linearized
trations of KA were normalized by the response to the EC
with the necessary restriction enzyme and then purified to serve as
concentration (100 M) immediately preceding the test concentration.
template for in vitro transcription. Briefly, in vitro cRNA transcripts
First, a response was recorded to 100 M KA, and after a 5-min
were prepared with the appropriate mMessage mMachine kit from
washout a response was recorded to a test dose of KA. After 5 min,
Ambion (Austin, TX). Transcription reactions were performed with 1
100 M KA was applied again to determine any residual effects from
g cDNA as template, an RNA polymerase (T3 or T7, depending on
the test dose of KA. If the response to 100 M KA after the test dose
clone), DTT, RNase inhibitor, dNTPs, and 32P. Nucleotide incorpo-
was Ն75% of the response to 100 M KA before the test dose, the
ration was evaluated by DEAE81 filter binding assays and a liquid
oocyte was tested further at other concentrations of KA. The resulting
scintillation counter. RNA was stored in DEPC water stocks at
concentration-response relationship was fitted with Eq. 2 (Luetje and
80°C, and aliquots were used for injection into the oocytes. The
accession numbers for the clones used in this study were GluR1(X17184), GluR2 (M85035), GluR3 (M85036), GluR6 (Z11548),
NMDAR1–1a (L19708), NMDAR2a (AF001423), and NMDAR2b
(U11419). Unless otherwise noted, we used the flop variants ofAMPA receptors in our experiments. Effect of Liϩ on neuronal GluR function
For the conventional two-electrode, voltage-clamp experiments,
Specific GluR subtypes were evaluated for their potential
oocytes were placed in a Warner Instruments (Hamden, CT) recording
modulation by Liϩ. Effects on GluR function were noted in the
chamber and perfused with frog Ringer solution [containing (in mM)
range of 5–115 mM Liϩ. For our standard agonist applications
115 NaCl, 2.5 KCl, 1.8 BaCl , and 10 HEPES, pH 7.3]. Ringer
we used 100 M KA for AMPA receptors, 100 M glutamate
solutions containing lithium (Liϩ) or rubidium were made by substi-
for KA receptors, and 100 M glutamate ϩ 10 M glycine for
tuting the ion for sodium. Osmolarity of different Ringer solutions
NMDA receptors. Under these conditions, KA/NMDA-recep-
was checked with a Precision Systems (Natick, MA) Osmette Aosmometer. A Warner Instruments Oocyte Clamp OC-725B and Fre-
tor currents appeared to be reduced in Liϩ solutions, whereas
quency Devices model 902 filter were interfaced with National In-
AMPA receptor currents were potentiated in Liϩ solutions.
struments (Austin, TX) LabVIEW software and a Macintosh com-
Subtype-selective differences in potentiation were observed
puter for data acquisition. Electrodes were fabricated from glass
within the AMPA receptor class and were investigated further
capillary tubes (KG-33) from Garner Glass (Claremont, CA) with a
with ionic variation in extracellular solutions, different ago-
DKI (Tujunga, CA) model 750 needle/pipette puller. Voltage elec-
nists at multiple concentrations, and chemical modulators.
trodes were filled with 3 M KCl and had resistances on the order of1–5 M⍀, whereas current electrodes were filled with 0.25 M CsCl,0.25 M CsF, and 100 mM EGTA (pH 7.3) and had resistances of
Permeability and conductance of neuronal GluRs to Liϩ
0.5–3 M⍀. Experiments were performed at room temperature, and the
Compared with their respective sodium controls, the ampli-
oocyte membrane was clamped at Ϫ50 mV. Currents were measured
tude of the KA-evoked current in Liϩ varied across subtype of
to the nearest nanoampere. At least three and usually four or moreoocytes were used for each measurement. Drugs were dissolved in
GluR. However, when extracellular sodium was replaced with
Ringer and applied by filling a 2.0-ml length of tubing at the end of
Liϩ, no shift in reversal potential was detected for any of the
the perfusion line. A discrete volume of agonist was thereby admin-
subtypes that were tested. For the AMPA-selective flop vari-
istered over a 10-s period. Some drug stocks were dissolved in DMSO
ants of GluRs, both inward and outward currents in Liϩ Ringer
Current-voltage relationships of various glutamate re-
ceptor (GluR) subtypes. A: current-voltage relationship a Xenopusoocyte expressing the ␣-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)-selective ionotropic GluR3 ϩ GluR2 in re-sponse to 100 M kainate (KA). B: bar graph of the currentrecorded in lithium (Liϩ) relative to Naϩ for GluR1, GluR1 ϩ R2,GluR3 ϩ GluR2, and GluR3 receptors with 100 M KA at 2different membrane voltages. Error bars represent SE. C: bar graphof the current recorded in Liϩ relative to Naϩ for GluR6Q andGluR6R receptors with 100 M glutamate at 2 different membranevoltages. D: bar graph of the current recorded in Liϩ relative toNaϩ for NMDAR1a ϩ NMDAR2a and NMDAR1a ϩ NMDAR2breceptors with 100 M glutamate ϩ 10 M glycine at 2 differentmembrane voltages.
were equal to or greater than the current in Naϩ Ringer. A
creased 134 Ϯ 26% above control (n ϭ 3) in the presence of
representative I-V relationship for GluR3 ϩ R2 is shown in
Liϩ (Fig. 2B). These results indicate that GluR2 is intrinsically
Fig. 1A. The increase of both inward and outward currents
capable of Liϩ modulation. However, further experiments fo-
without a shift in the reversal potential suggests an effect on the
cused on the GluR3 subtype because it was most sensitive to
rather than an increase in single channel conductance
Liϩ potentiation and easily formed homomeric receptors.
with Liϩ. In general, the inward and outward currents ofvarious AMPA receptors were potentiated by Liϩ, rangingfrom 40 to 200% above control (Fig. 1B). However, the current
Specificity of the effect of Liϩ on GluR currents
evoked by 100 M KA in Liϩ Ringer through GluR1 was notsignificantly increased. In contrast, under these conditions the
To test the specificity of Liϩ effect on GluR currents, we
current in Liϩ Ringer was decreased by 40 – 60% for KA and
performed similar experiments in rubidium-based Ringer. Raw
NMDA receptors (Fig. 1, C and D). Because GluR1 ϩ R2
waveforms recorded from a GluR3-expressing oocyte in Naϩ,
receptor currents were potentiated but GluR1 currents were not
Rbϩ, and Liϩ Ringer are shown in Fig. 3A. Rubidium produced
(Fig. 1B), we investigated the role of the GluR2 subunit in Liϩ
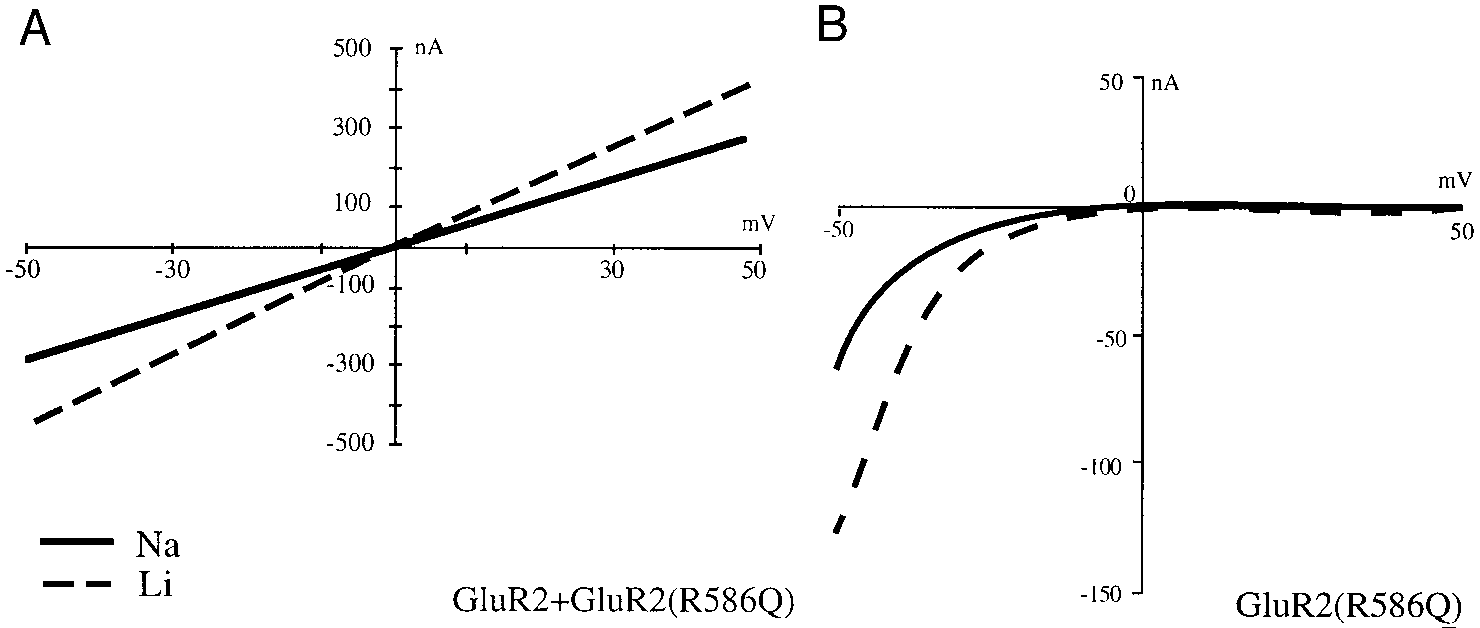
a mean increase in current of 56 Ϯ 4.6% (n ϭ 4) but corre-
potentiation. Wild-type GluR2 alone does not function well in
spondingly shifted the reversal potential 6.60 Ϯ 1.03 mV (n ϭ
oocytes. Therefore for these experiments we expressed wild-
7) for GluR3 (Fig. 3A, inset) and 4.05 Ϯ 0.46 mV (n ϭ 10) for
type GluR2 with a Q/R site mutant, GluR2(R586Q), as well as
GluR3 ϩ R2 (Fig. 4, A and B). There is no potentiation of
the mutant alone. The inward and outward currents of hetero-
outward current by Rbϩ for GluR3 (Fig. 3A, inset) or for
meric GluR2 ϩ GluR2(R586Q) receptors were increased in
GluR3 ϩ R2 (Fig. 4B). By using the shift in reversal potentials
Liϩ by 74 Ϯ 7.9% (n ϭ 6) and 72 Ϯ 10% (n ϭ 6), respectively
and solving for the ratio P /P
(Fig. 2A). The conductance of the mutant homomer was in-
of 1.30 for GluR3 and 1.17 for GluR3 ϩ R2.
Q/R site mutant heteromer and homomer. A:current-voltage relationship from a Xenopusoocyte expressing the AMPA-selective iono-tropic GluR2 ϩ GluR2(R586Q) in response to100 M KA. B: GluR2(R586Q).
LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
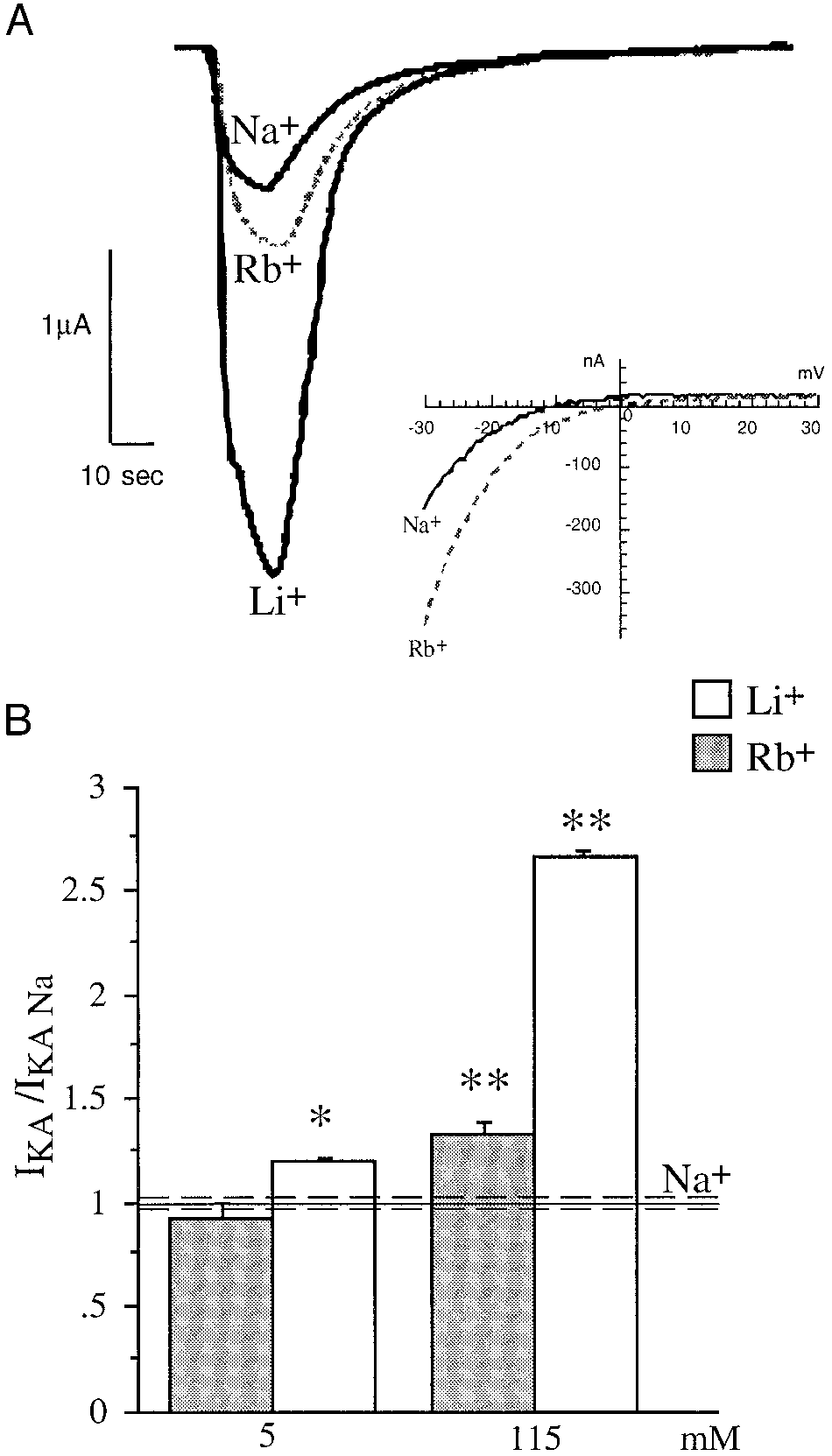
Effects of Rbϩ on GluR3 ϩ GluR2. A: mean data for the reversal
potential shift of GluR3 and GluR3 ϩ R2 caused by Rbϩ. B: I-V relationshipfor GluR3 ϩ GluR2 in Naϩ and Rbϩ. Note the rightward shift in reversalpotential and that the outward current is not affected as in Fig. 1A. The agonistused was 100 M KA.
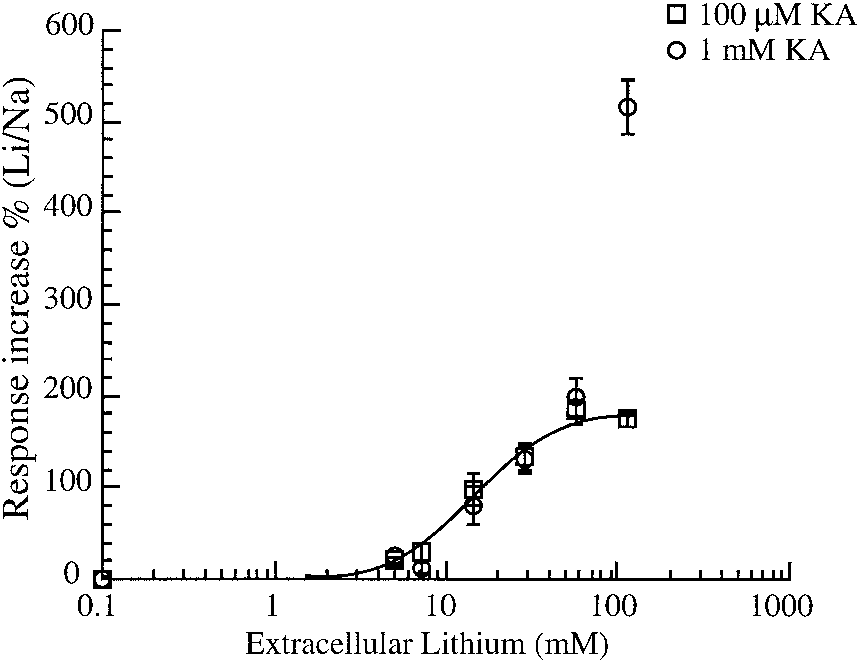
Liϩ, 20 mM sucrose). The presence of 10 mM Naϩ in 115 mMLiϩ Ringer did not significantly reduce the response to 1 mMKA (data not shown). These results indicate that the extremepotentiation reported for 1 mM KA responses obtained in thepresence of 115 mM Liϩ was not due to a specific effectassociated with the removal of sodium. Note that when 1 mMGlu was used as the agonist the GluR3 current displayed anincreased threshold for potentiation by Liϩ that did not appearto saturate and reached a maximum potentiation of 357% with100% mole fraction of Liϩ (Fig. 7B).
It is interesting to note the receptor sensitivity to potentiation
by concentrations of Liϩ that approach levels used for treat-ment of bipolar disorder. We observed an increase of 21 Ϯ 3%(n ϭ 10, P Ͻ 0.001) for GluR3 and 26 Ϯ 3% (n ϭ 6, P Ͻ 0.05)for GluR3 ϩ R2 in response to 100 M KA at 5 mM extra-cellular Liϩ. If the potentiation was linearly dependent onextracellular Liϩ concentration, at 5 mM extracellular Liϩ(1/23 of 115 mM) one would expect that the potentiation wouldbe ϳ1/23 of the maximum potentiation observed with 100 MKA and 115 mM Liϩ (or 7– 8%). This observation furthersupports the hypothesis that the effects of Liϩ are most likelydue to effects on P
response waveforms recorded in 115 mM Naϩ, 115 mM Liϩ,and 5 mM Liϩ from GluR3-expressing oocytes are shown in
Comparison of the effects of Rbϩ and Liϩ. A: response waveforms
Fig. 6, A–C. Rubidium has no effect on GluR3 current at 5 mM
evoked with 100 M KA from a GluR3 expressing oocyte in Naϩ, Rbϩ, andLiϩ Ringer solutions. The response of GluR3 in Naϩ Ringer is the smallest of
extracellular concentration in contrast to Liϩ (Figs. 3B and 6).
the 3 solutions tested. The response of GluR3 in Rbϩ Ringer is ϳ56% larger
Potentiation by Liϩ is readily reversible at both high and low
than in Naϩ and is approximately 170% larger in Liϩ Ringer than in Naϩ. The
concentrations on washout of the Liϩ Ringer. In Fig. 6D, an
inset shows a representative I-V for GluR3 in Naϩ and Rbϩ. The reversalpotential is shifted 6.60 Ϯ 1.03 mV in the positive direction. B: comparison ofthe potentiation caused by 2 different concentrations of Liϩ (open bar) andRbϩ (shaded bar), relative to Naϩ (solid line; SE, dashed line). Asterisksindicate significant differences compared with Naϩ. * P Ͻ 0.001, ** P Ͻ0.0001. Concentration dependence of extracellular Liϩ
We wished to determine if the magnitude of Liϩ potentiation
of GluR3 response varies linearly with external Liϩ concen-tration. We noted that when 100 M KA is used as the agonistan increase of the extracellular Liϩ concentration causes acurrent potentiation that could be fit by Eq. 2 with an EC
14.85 Ϯ 1.71 mM (n Ն 4) (Fig. 5). However, when saturatingconcentrations (1 mM) of KA were used we saw a furtheranomalous increase in potentiation at the highest extracellularLiϩ concentration. To test if this peculiar increase with 1 mMKA in 100% Liϩ (115 mM Liϩ, 0 mM Naϩ) could be asso-
Effect of extracellular [Liϩ] titration on GluR3 response potentia-
ciated with the total absence of sodium we performed experi-
tion. The percent increase in response caused by Liϩ relative to Naϩ Ringer isplotted vs. extracellular [Liϩ] for GluR3 expressing oocytes. Two different
ments at the same Liϩ concentration but with added Naϩ (115
agonist concentrations were used; 100 M KA (Ⅺ) and 1 mM KA (E). Each
mM Liϩ, 10 mM Naϩ) as well as an osmotic control (115 mM
point represents the means Ϯ SE of 4 –11 oocytes.
therapeutic window. A: depicted here are re-sponses to 100 M KA in Naϩ Ringer fromoocytes expressing GluR3 channels. B: first 2responses are recorded in Naϩ Ringer, the next2 in Ringer with 115 mM Liϩ, and the finalresponse is a washout back to Naϩ Ringer. C:paradigm is the same as in B except the thirdand fourth responses are recorded in 110 mMNaϩ/5 mM Liϩ. D: I-V relationship for a Xe-nopus oocyte expressing GluR3 ϩ GluR2 re-corded in 110 mM Naϩ/5 mM Liϩ showingpotentiation of both inward and outward cur-rent. I-V relationship of a GluR3 ϩ R2-expressing oocyte recorded
Because flip/flop variants were reported to vary in their
in 5 mM Liϩ showed an increase of both inward and outward
intrinsic desensitization, we set out to determine the interaction
currents analogous to that shown in Fig. 1A.
of the flip/flop domain with Liϩ potentiation. We investigatedGluR3 flip currents for their capacity to be modulated by Liϩ
Liϩ effects on GluR agonist potency
at saturating agonist concentrations in the presence and ab-sence of CTZ and Liϩ (Fig. 8, E and F). We found that Liϩ
We examined the potency and efficacy of KA in the pres-
potentiated the current evoked from GluR3 flip channels less
ence and absence of Liϩ. The concentration-response relation-ship determined for GluR3 in Naϩ Ringer yielded a Hill slopeof 1.14 Ϯ 0.07 and an EC
Liϩ Ringer, the concentration-response relationship yielded aHill slope of 2.19 Ϯ 0.56 and an EC
7A, Œ). Liϩ increased the maximal attainable response by
Ͼ500% and increased the EC by 60% compared with so-
dium. The total percent increase caused by Liϩ for glutamateand KA at low and high concentrations relative to the sameagonist and concentration in Naϩ is presented in Fig. 7B. GluR1 does not show the same magnitude of increase com-pared with GluR3 with either KA or glutamate at saturatingconcentrations. Liϩ effects and desensitization
We examined the effect of Liϩ when desensitization of
GluR3 channels was reduced with the compound CTZ (Fig. 8). With 100 M KA or glutamate as the agonist, both Liϩ and100 M CTZ potentiate GluR3 responses to the same extent. There was no further increase when the treatments were com-bined (Fig. 8, A and B). With 1 mM KA as the agonist, thecombination of 100 M CTZ and Liϩ substitution produced acurrent increase that was 30% larger than the increase CTZproduced in Naϩ Ringer (Fig. 8C, P Ͻ 0.05, unpaired t-test).
Concentration-response relationship. A: concentration-response re-
At a high glutamate concentration (1 mM), the combination of
lationship of KA for GluR3 in Naϩ and Liϩ Ringer solution. All responses
100 M CTZ and Liϩ substitution produced a current increase
were normalized to the response of 100 M KA in Naϩ Ringer. B: percent
that was 353 Ϯ 54% larger than the increase CTZ produced in
increase in response caused by Liϩ with different agonists and agonist con-
Naϩ Ringer (Fig. 8D, P Ͻ 0.001, unpaired t-test). Although
centrations. For 100 M KA the increase was 220 Ϯ 14% (n ϭ 19), for 1 mM
CTZ potentiated GluR1 currents in sodium, there was no
KA the increase was 518 Ϯ 30% (n ϭ 19), for 100 M glutamate the increasewas 237 Ϯ 29% (n ϭ 5), and for 1 mM glutamate the increase was 357 Ϯ 54%
apparent interaction between the combination of CTZ with Liϩ
(n ϭ 11). The increase is presented normalized to the response to the respective
at either agonist concentration (data not shown).
LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
than from GluR3 flop. Potentiation of responses to 1 mM KA
(Fig. 8F, P Ͻ 0.05, unpaired t-test). This reduction in GluR3
and 1 mM Glu was only 19 and 67% of the potentiation
flip current when CTZ and Liϩ are combined was also seen
obtained with GluR3 flop. We also examined the effect of Liϩ
when lower agonist concentrations were used (100 M Glu),
when desensitization of GluR3 flip channels was reduced with
indicating that this effect is not specific to high agonist con-
the compound CTZ (Fig. 8, E and F). In contrast to the results
centrations (n ϭ 3, P Ͻ 0.05, data not shown).
obtained with GluR3 flop, Liϩ actually decreased 1 mM KA ϩ100 M CTZ-evoked GluR3 flip currents by 19% (n ϭ 6)
compared with the 1 mM KA ϩ 100 M CTZ-evoked currentsin Naϩ Ringer (n ϭ 6) (Fig. 8E, P Ͻ 0.05, unpaired t-test).
We characterized the subtype-selective potentiation of
Similarly, Liϩ decreased 1 mM Glu ϩ 100 M CTZ-evoked
GluR3 by Liϩ through an evaluation of Liϩ effects with
GluR3 flip currents by 35% (n ϭ 11) compared with the 1 mM
agonist concentration, mole fraction of extracellular Liϩ, and
Glu ϩ 100 M CTZ-evoked currents in Naϩ Ringer (n ϭ 11)
compounds affecting receptor desensitization. Our results per-mit us to propose that the effect of Liϩ is to modify desensi-tization in a manner that depends on the flop domain.
Liϩ, rubidium, and sodium are all monovalent cations in
group I of the periodic table, but only Liϩ is used for thetreatment of the mental illness bipolar disorder. Studies involv-ing Liϩ inhibition of second-messenger systems often userubidium to confirm the specificity of the inhibition by Liϩ(Ebstein et al. 1980). We therefore compared GluR3 currents inLiϩ-, Naϩ-, and Rbϩ-based Ringer. Although we observed apotentiation of the responses in both Rbϩ and Liϩ Ringercompared with Naϩ, the potentiation in Rbϩ was qualitativelydifferent than in Liϩ because it coincided with a shift of thereversal potential in the positive direction and was not ob-served on outward currents. These data suggest that the effectsof Rbϩ were largely due to an increased permeability of Rbϩthrough the channel.
The magnitude of Liϩ potentiation increased with the con-
centration of extracellular Liϩ independently of agonist con-centration except at saturating agonist concentration in thepresence of 115 mM Liϩ. One potential explanation for thisobservation is that there are two processes that contribute toLiϩ potentiation of GluR current. One process, which is inde-pendent of agonist concentration, may predominate at lowerLiϩ concentrations. At very high Liϩ concentrations, a secondform of potentiation may manifest that selectively enhancesresponses to high, potentially desensitizing concentrations ofagonists.
The results of our concentration-response experiments fur-
ther suggest that Liϩ caused a dramatic change in the apparentefficacy of the agonist. A raised maximal response suggests anincrease in the probability of a channel being open (P ) or an
increase in the single-channel conductance (␥). However, theeffects of Liϩ on outward current, where it is not the primarycharge carrier, favors the interpretation that there is an alter-
Interactions among cyclothiazide (CTZ), agonist, and Liϩ. Shaded
ation in the percentage of time that channels are open.
bars represent mean current potentiation in Liϩ relative to Naϩ (dashed line).
CTZ was reported to potentiate AMPA receptor currents by
Open bars represent mean current potentiation by 100 M CTZ relative to Naϩ
reducing receptor desensitization (Partin et al. 1996). Because
(dashed line). Striped bars represent mean current potentiation in Liϩ ϩ100
Liϩ effects were greatest under desensitizing conditions, we
M CTZ relative to Naϩ (dashed line). Error bars represent SE. A: GluR3 flop
hypothesized that the effect of Liϩ might also be to reduce
responses to 100 M KA in 115 mM Liϩ (n ϭ 13), 115 mM Naϩ ϩ 100 MCTZ (n ϭ 13), and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 14). B: GluR3 flop
desensitization, and we sought to determine if CTZ and Liϩ
responses to 100 M glutamate in 115 mM Liϩ (n ϭ 7), 115 mM Naϩ ϩ 100
acted through similar or different mechanisms. If Liϩ and CTZ
M CTZ (n ϭ 7), and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 8). C: GluR3 flop
work through distinct mechanisms, the potentiation that each
responses to 1 mM KA in 115 mM Liϩ (n ϭ 9), 115 mM Naϩ ϩ 100 M CTZ
causes individually might be additive when they are applied in
(n ϭ 9), and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 7). D: GluR3 flop responsesto 1 mM glutamate in 115 mM Liϩ (n ϭ 4), 115 mM Naϩ ϩ 100 M CTZ
combination. Our results indeed suggest that, at high concen-
(n ϭ 4), and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 4). E: GluR3 flip responses
trations of KA, Liϩ and CTZ do not act entirely through the
to 1 mM KA in 115 mM Liϩ (n ϭ 6), 115 mM Naϩ ϩ 100 M CTZ (n ϭ 6),
same mechanism. Moreover, when a high concentration of
and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 6). F: GluR3 flip responses to 1 mM
glutamate (1 mM) was used, the effects of Liϩ and CTZ
glutamate in 115 mM Liϩ (n ϭ 11), 115 mM Naϩ ϩ 100 M CTZ (n ϭ 11),
appeared to be completely additive for GluR3. These observa-
and 115 mM Liϩ ϩ 100 M CTZ (n ϭ 11). Asterisks indicate significantdifferences compared with Naϩ ϩ CTZ. * P Ͻ 0.05, ** P Ͻ 0.001.
tions are consistent with the idea that Liϩ produces potentia-
tion of current in GluR3 by modulating the amount of receptor
BLISS, T.V.P. AND LYNCH, M. A. Long-term potentiation of synaptic transmis-
desensitization and that desensitization of the channel can be
sion in the hippocampus: properties and mechanisms. In: Long-term Poten-
influenced by specific agonists. Desensitization is also con-
tiation: From Biophysics to Behavior. New York: Liss, 1988, p. 3–72.
COLLINGRIDGE, G. L. AND BLISS, T.V.P. NMDA receptors: their role in long-
trolled by the presence of specific protein domains, as in the
term potentiation. Trends Neurosci. 10: 288 –293, 1987.
alternative splice variants, flip and flop, in AMPA receptors.
EBSTEIN, R. P., HERMONI, M., AND BELMAKER, R. H. The effect of lithium on
Flop receptors are thought to desensitize more than flip vari-
noradrenaline-induced cyclic AMP accumulation in rat brain: inhibition
ants. Indeed, it appears that flop receptor currents are also
after chronic treatment and absence of supersensitivity. J Pharmacol. Exp.
potentiated to a greater degree by Liϩ than are flip receptor
currents. The specificity of the Liϩ effect for the flop domain
HOLLMANN, M., HARTLEY, M., AND HEINEMANN, S. Ca2ϩ permeability of
KA-AMPA-gated glutamate receptor channels depends on subunit compo-
is further supported by the observation that, under conditions
sition. Science 252: 851– 853, 1991.
of the additive CTZ effect for flop variants, there was an
KARKANIAS, N. B., KABAKOV, A. Y., LENOX, R. H., AND PAPKE, R. L. Synapse-
antagonism of the CTZ effect in flip variants.
specific accumulation of lithium in intracellular microdomains: a model for
The inconsistent interaction of Liϩ with CTZ suggests a
uncoupling coincidence detection in the brain. Synapse 28: 271–279, 1998.
novel mechanism for this effect, although with a similar re-
LOMELI, H., MOSBACHER, J., MELCHER, T., HOGER, T., GEIGER, JR., KUNER, T.,
quirement for the flop domain. The modulation of desensitiza-
MONYER, H., HIGUCHI, M., BACH, A., AND SEEBURG, P. H. Control of kineticproperties of AMPA receptor channels by nuclear RNA editing. Science
tion could involve alterations of rate constants into or out of the
desensitized state as well as alterations of rate constants lead-
LUETJE, C. W. AND PATRICK, J. Both ␣- and -subunits contribute to the agonist
ing toward the desensitized state. Our data suggest that further
sensitivity of neuronal nicotinic acetylcholine receptors. J. Neurosci. 11:
detailed studies of desensitization may exploit the use of Liϩ as
a tool to dissect the mechanisms of desensitization.
MONYER, H., SPRENGEL, R., SCHOEPFER, R., HERB, A., HIGUCHI, M., LOMELI,
H., BURNASHEV, N., SAKMANN, B., AND SEEBURG, P. H. Heteromeric NMDA
The authors thank Drs. Michael Hollmann, Jane Sullivan, Steve Heinemann,
receptors: molecular and functional distinction of subtypes. Science 256:
and Jim Boulter for the glutamate receptor (GluR) subunit genes and Dr. R.
Dingledine for providing the mutant GluR2 channel cDNA. We thank Dr.
MOSBACHER, J., SCHOEPFER, R., MONYER, H., BURNASHEV, N., SEEBURG, P.,
Robert Lenox for helpful discussion.
AND RUPPERSBERG, J. A molecular determinant for submillisecond desensi-
This work was supported by predoctoral fellowships to N. B. Karkanias
tization in glutamate receptors. Science 266: 1059 –1062, 1994.
from the University of Florida Center for Neurobiology and Behavior and the
PARTIN, K. M., FLECK, M. W., AND MAYER, M. L. AMPA receptor flip/flop
University of Florida Division of Sponsored Research. Technical assistance
mutants affecting deactivation, desensitization, and modulation by cy-
clothiazide, aniracetam, and thiocyanate. J. Neurosci. 16: 6634 – 6647, 1996.
Address for reprint requests: R. L. Papke, Depts. Of Pharmacology &
PARTIN, K. M., PATNEAU, D. K., WINTERS, C. A., MAYER, M. L., AND BUO-
Therapeutics and Neuroscience, University of Florida Medical College, J. H.
NANNO, A. Selective modulation of desensitization at AMPA versus kainate
Miller Health Center, Box 100267, Gainesville, FL 32610-0267.
receptors by cyclothiazide and concanavalin A. Neuron 11: 1069 –1082,1993.
Received 14 September 1998; accepted in final form 15 December 1998.
SOMMER, B., KEIN¨ANEN, K., VERDOORN, T. A., WISDEN, W., BURNASHEV, N.,
HERB, A., KOHLER, M., TAKAGI, T., SAKMANN, B., AND SEEBURG, P. H. Flip
and flop: a cell-specific functional switch in glutamate-operated channels ofthe CNS. Science 249: 1580 –1585, 1990.
BARBOUR, B., KELLER, B. U., LLANO, I., AND MARTY, A. I. Prolonged presence
VYKLICKY, J. L., PATNEAU, D. K., AND MAYER, M. L. Modulation of excitatory
of glutamate during excitatory synaptic transmission to cerebellar Purkinje
synaptic transmission by drugs that reduce desensitization at AMPA/kainate
cells. Neuron 12: 1331–1343, 1994.
receptors. Neuron 7: 971–984, 1991.
BLISS, T.V.P. AND GARDNER-MEDWIN, A. R. Long-lasting potentiation of
WONG, L. A. AND MAYER, M. L. Differential modulation by cyclothiazide and
synaptic transmission in the dentate area of the unanaesthetized rabbit
concanavalin A of desensitization at native alpha-amino-3-hydroxy-5-
following stimulation of the perforant path. J. Physiol. (Lond.) 232: 357–
methyl-4-isoxazolepropionic acid- and kainate-preferring glutamate recep-
tors. Mol. Pharmacol. 44: 504 –510, 1993.
TREATING Based on Practice Guideline for the Treatment of Patients With Obsessive-Compulsive Disorder,originally published in July 2007. A guideline watch, summarizing significant developments in the scientific literature since publication of this guideline, may be available in the Psychiatric Practice section of the APA web site at www.psych.org. American Psychiatric Association Steer
klüger, schneller – Gentech möglich! Basler Appell gegen Gentechnologie Editorial War es vor einigen Jahren der menschliche «Gen-Pool», der verbessert werden sollte,so arbeiten heute Forschungseinrichtungen in aller Welt daran, uns auf individueller Human Enhancement – Die Verbesserung des MenschenEbene zu «optimieren». Körperliche Leistungsfähigkeit, mentales
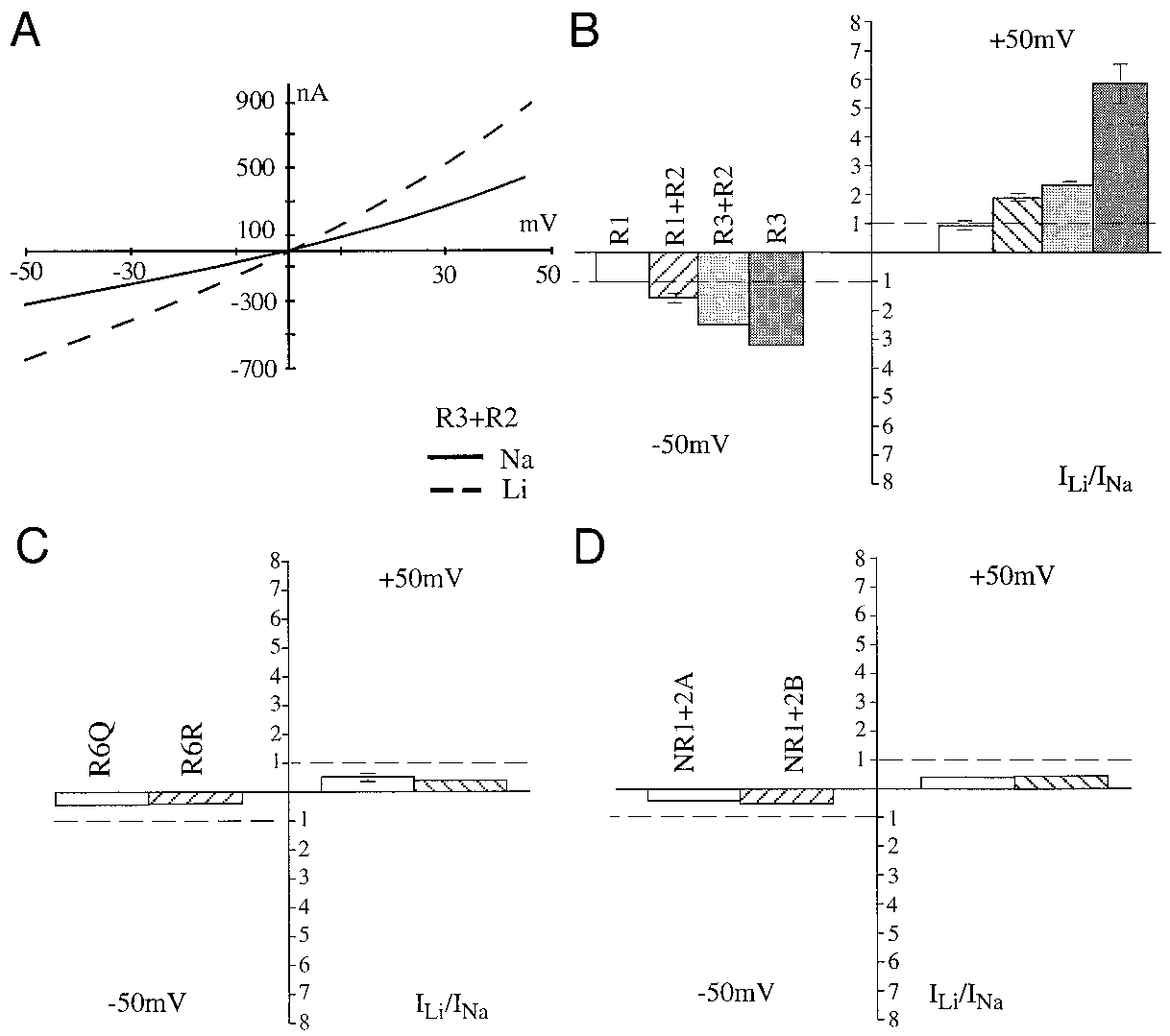
 Current-voltage relationships of various glutamate re-
ceptor (GluR) subtypes. A: current-voltage relationship a Xenopusoocyte expressing the ␣-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)-selective ionotropic GluR3 ϩ GluR2 in re-sponse to 100 M kainate (KA). B: bar graph of the currentrecorded in lithium (Liϩ) relative to Naϩ for GluR1, GluR1 ϩ R2,GluR3 ϩ GluR2, and GluR3 receptors with 100 M KA at 2different membrane voltages. Error bars represent SE. C: bar graphof the current recorded in Liϩ relative to Naϩ for GluR6Q andGluR6R receptors with 100 M glutamate at 2 different membranevoltages. D: bar graph of the current recorded in Liϩ relative toNaϩ for NMDAR1a ϩ NMDAR2a and NMDAR1a ϩ NMDAR2breceptors with 100 M glutamate ϩ 10 M glycine at 2 differentmembrane voltages.
Current-voltage relationships of various glutamate re-
ceptor (GluR) subtypes. A: current-voltage relationship a Xenopusoocyte expressing the ␣-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA)-selective ionotropic GluR3 ϩ GluR2 in re-sponse to 100 M kainate (KA). B: bar graph of the currentrecorded in lithium (Liϩ) relative to Naϩ for GluR1, GluR1 ϩ R2,GluR3 ϩ GluR2, and GluR3 receptors with 100 M KA at 2different membrane voltages. Error bars represent SE. C: bar graphof the current recorded in Liϩ relative to Naϩ for GluR6Q andGluR6R receptors with 100 M glutamate at 2 different membranevoltages. D: bar graph of the current recorded in Liϩ relative toNaϩ for NMDAR1a ϩ NMDAR2a and NMDAR1a ϩ NMDAR2breceptors with 100 M glutamate ϩ 10 M glycine at 2 differentmembrane voltages.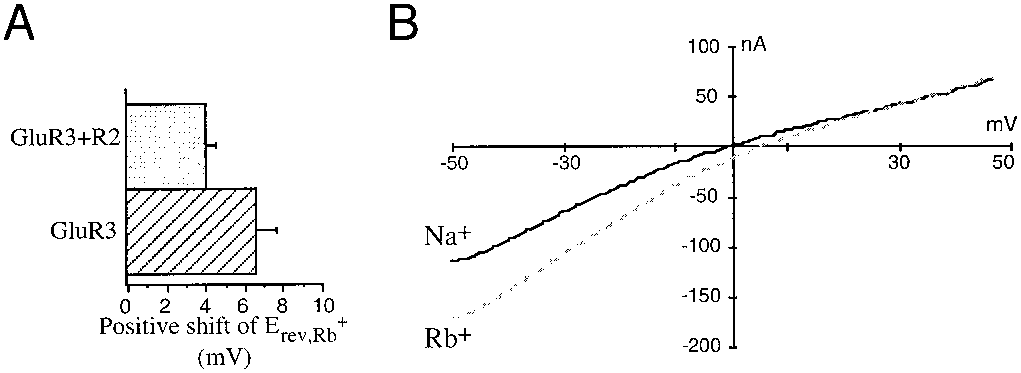
 LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
Effects of Rbϩ on GluR3 ϩ GluR2. A: mean data for the reversal
potential shift of GluR3 and GluR3 ϩ R2 caused by Rbϩ. B: I-V relationshipfor GluR3 ϩ GluR2 in Naϩ and Rbϩ. Note the rightward shift in reversalpotential and that the outward current is not affected as in Fig. 1A. The agonistused was 100 M KA.
LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
Effects of Rbϩ on GluR3 ϩ GluR2. A: mean data for the reversal
potential shift of GluR3 and GluR3 ϩ R2 caused by Rbϩ. B: I-V relationshipfor GluR3 ϩ GluR2 in Naϩ and Rbϩ. Note the rightward shift in reversalpotential and that the outward current is not affected as in Fig. 1A. The agonistused was 100 M KA.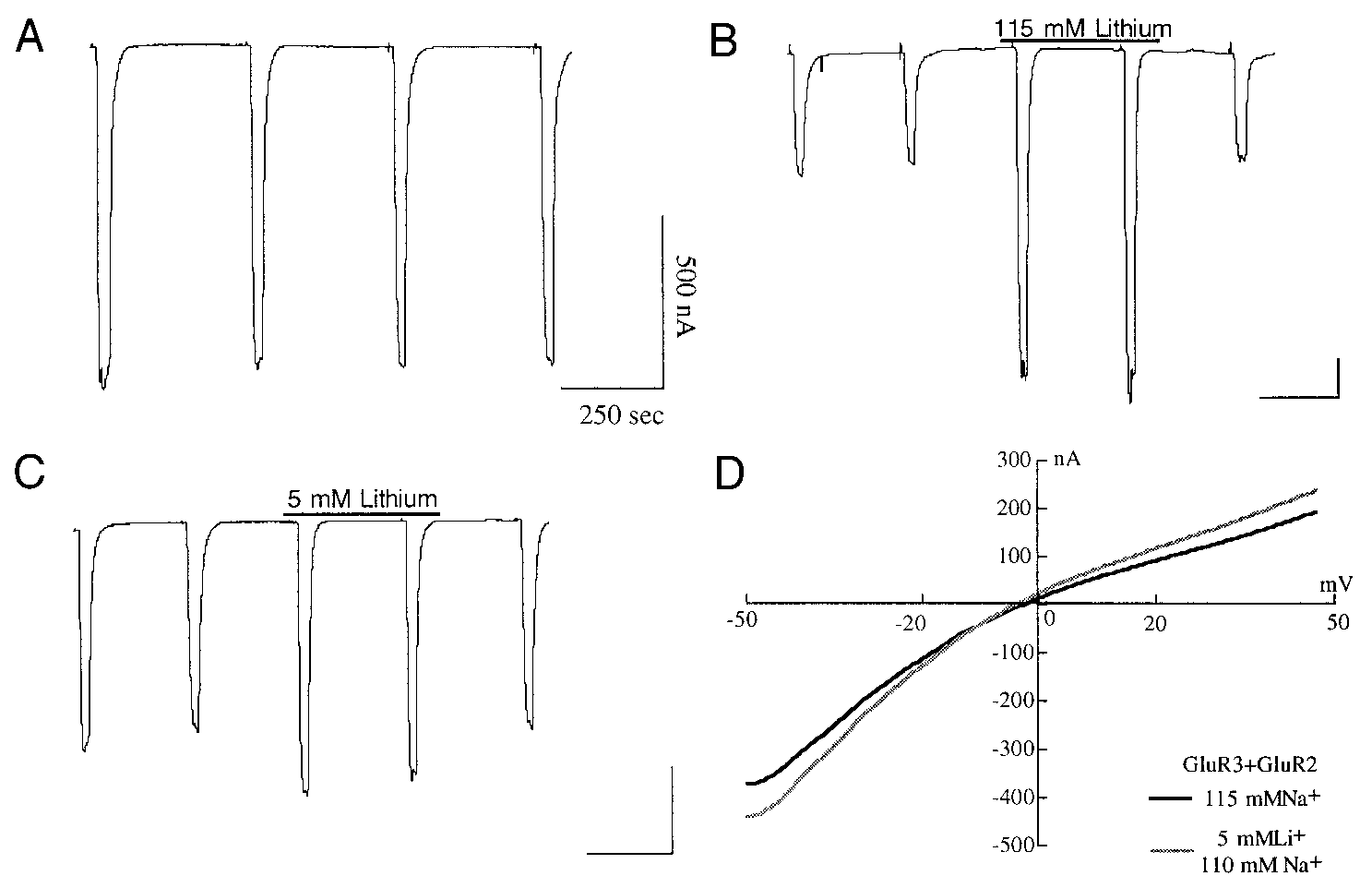
 therapeutic window. A: depicted here are re-sponses to 100 M KA in Naϩ Ringer fromoocytes expressing GluR3 channels. B: first 2responses are recorded in Naϩ Ringer, the next2 in Ringer with 115 mM Liϩ, and the finalresponse is a washout back to Naϩ Ringer. C:paradigm is the same as in B except the thirdand fourth responses are recorded in 110 mMNaϩ/5 mM Liϩ. D: I-V relationship for a Xe-nopus oocyte expressing GluR3 ϩ GluR2 re-corded in 110 mM Naϩ/5 mM Liϩ showingpotentiation of both inward and outward cur-rent.
therapeutic window. A: depicted here are re-sponses to 100 M KA in Naϩ Ringer fromoocytes expressing GluR3 channels. B: first 2responses are recorded in Naϩ Ringer, the next2 in Ringer with 115 mM Liϩ, and the finalresponse is a washout back to Naϩ Ringer. C:paradigm is the same as in B except the thirdand fourth responses are recorded in 110 mMNaϩ/5 mM Liϩ. D: I-V relationship for a Xe-nopus oocyte expressing GluR3 ϩ GluR2 re-corded in 110 mM Naϩ/5 mM Liϩ showingpotentiation of both inward and outward cur-rent. LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
than from GluR3 flop. Potentiation of responses to 1 mM KA
(Fig. 8F, P Ͻ 0.05, unpaired t-test). This reduction in GluR3
and 1 mM Glu was only 19 and 67% of the potentiation
flip current when CTZ and Liϩ are combined was also seen
obtained with GluR3 flop. We also examined the effect of Liϩ
when lower agonist concentrations were used (100 M Glu),
when desensitization of GluR3 flip channels was reduced with
indicating that this effect is not specific to high agonist con-
the compound CTZ (Fig. 8, E and F). In contrast to the results
centrations (n ϭ 3, P Ͻ 0.05, data not shown).
LITHIUM AFFECTS GLUTAMATE RECEPTOR FUNCTION
than from GluR3 flop. Potentiation of responses to 1 mM KA
(Fig. 8F, P Ͻ 0.05, unpaired t-test). This reduction in GluR3
and 1 mM Glu was only 19 and 67% of the potentiation
flip current when CTZ and Liϩ are combined was also seen
obtained with GluR3 flop. We also examined the effect of Liϩ
when lower agonist concentrations were used (100 M Glu),
when desensitization of GluR3 flip channels was reduced with
indicating that this effect is not specific to high agonist con-
the compound CTZ (Fig. 8, E and F). In contrast to the results
centrations (n ϭ 3, P Ͻ 0.05, data not shown).