La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Nugi-zentrum.de
Rafael Gómez-Lus
Department of Microbiology, Hospital Clínico
Received 25 June 1998Accepted 25 July 1998
Summary Bacterial resistance to antibiotics is often plasmid-mediated and the associated genes encoded by transposable elements. These elements play a central role in evolution by providing mechanisms for the generation of diversity and, in conjunction with DNA transfer systems, for the dissemination of resistances to other bacteria. At the University Hospital of Zaragoza, extensive efforts have been made to define both the dissemination and evolution of antibiotic resistance by studying the transferable R plasmids and transposable elements. Here we describe the research on bacterial resistance to antibiotics in which many authors listed in the references have participated. The aspects of bacterial resistance dealt with are: (i) transferable resistance mediated by R plasmids in Gram- negative bacteria, (ii) R plasmid-mediated resistance to apramycin and hygromycin
in clinical strains, (iii) the transposon Tn1696 and the integron In4, (iv) expression of
Department of Microbiology. Hospital Clínico Universitario.
Escherichia coli resistance genes in Haemophilus influenzae, (v) aminoglycoside-
modifying-enzymes in the genus Mycobacterium with no relation to resistance, and (vi)
macrolide-resistance and new mechanisms developed by Gram-positive bacteria
Tel.: +34-976761759. Fax: +34-976761693. E-mail: gomezlus@posta.unizar.es
characterized the plasmid pUZ1 (initially R1033) from a strain
of Pseudomonas aeruginosa coming from the Regional Centerfor Traumatology, near the University Hospital of Zaragoza. The
Bacterial resistance to antibiotics in Gram-negative bacilli is
plasmid had a size of 68 kb. Its incompatibility group was P (IncP)
most commonly mediated by R plasmids and by genes carried
and it was transferable to Escherichia coli J62 by conjugation.
by transposons (Tn) and integrons (In). In Gram-positive cocci
The plasmid coded for resistance to ampicillin, tetracycline,
the conjugative chromosomal transposons are fundamental for
gentamicin, kanamycin, streptomycin, chloramphenicol,
antibiotic resistance. They are characterized by their related
sulfamides and mercuric chloride [19]. The donor as well as
features to R plasmids, bacteriophages and the classic
the recipient strain coded for the enzymes TEM-1, AAC(3)-I,
transposons (for instance, Tn5 and Tn10). All these genetic
APH(3’)-I, and ANT(3’’) [9]. Plasmid pUZ1 contained two
elements play a central role in bacterial evolution, providing
transposons, the already known Tn3, which carries the blaTEM-1
mechanisms to generate diversity and having DNA transfer
gene, and a new transposon, Tn1696 [17], which bears the genes
systems able to disseminate sequences among other bacteria. aacC1, aadA1, sul1 and mer, which will be discussed later.
Good examples of those phenomena are the participation of
Four years later, we isolated different strains of
transposons in the rapid spread of antibiotic resistance among
enterobacteria and of P. aeruginosa (Table 1), which contained
bacterial populations—above all in those causing nosocomial
also plasmids of the IncP group, of the same size, with identical
infections—and the role of integrons in the maintenance and
restriction patterns and enzymes as pUZ1 [6, 15]. These findings
spread of the sul1 gene in enterobacteria.
supported the hypothesis that R plasmids spread to proximalareas, with the advantage of being able to propagate betweenthe Enterobacteriaceae and Pseudomonadaceae, which is
In 1976 we detected a 73 kb plasmid which belonged to the
group IncM and, thus, only transferable to enterobacteria. The
At the University Hospital of Zaragoza and other hospital centres
plasmid conferred resistance to ampicillin, tetracycline, gentamicin,
in Zaragoza, research has been carried out regarding the evolution
tobramycin and dibekacin and produced the enzymes TEM-1 and
and spread of antibiotic resistance over the last 30 years. We have
AAC(3)-V [14]. Following this, numerous strains of enterobacteria
studied conjugative R plasmids (tra+), transposable elements and
were isolated which also carried plasmids from the IncM group,
the biochemical mechanisms of resistance. In 1974 we
with identical properties, genotypes and phenotypes (Table 2).
In one of the strains of Proteus vulgaris (strain 18182) two
unit initiated by the blaTEM-1 gene promoter, which is
conjugative plasmids were detected. One was from the IncM
responsible for the mobilization of one of the IS140.
group (pUZ3a) and the other from the IncP group (pUZ3b). The
The capacity for transposition between replicons
pattern of resistance transferred corresponds to that described in
containing the bla genes is noteworthy, and it is reflected in
Tables 2 and 1, respectively. In addition, as a result of the
the numerous plasmid incompatibility groups, in which the
epidemiological monitoring of the strains carrying R plasmids,
bla genes have been detected: 18 for β-lactamase TEM-1, 7
we found that certain plasmids which were initially transferable
for TEM-2, 3 for OXA-1, 4 for OXA-2, and 2 for OXA-3.
(tra+) later lost this capacity (tra–). We also found plasmid loss or
We have already referred to part of our experiments involving
recombination of plasmid genes with the chromosome, which
the blaTEM-1 genes isolated from plasmids in the groups IncP
and IncM. In France, the gene blaTEM-3 has also been
A case to point out was a transconjugant of E. coli characterized in R plasmids from the IncM group. This gene
(strain 3644) which had lost the plasmid it was carrying but
codes for plasmid β-lactamases of extended spectrum.
had conserved its pattern of resistance. The alternative strategy
Previously, in the same country, an IncC plasmid had been
was to use the plasmid pUZ8, lacking transposable elements,
described which carried a blaTEM-4 gene coding for an
and introduce it into the aforementioned transconjugant of E.
extended β-lactamase spectrum, named TEM-4. coli J62 (F– his lac nalR pro trp). Through conjugation withE. coli J53 (F– met pro rifR), a plasmid (pUZ3644) was obtainedwhich contained a 27 kb fragment, carrying the blaaacC5 genes, flanked by two copies of an insertion sequence,
IS140. In this segment, the composite transposon Tn2922 was
characterized. Tn2922 is a co-integrate capable of transposingas a unit and formed by the fusion of the genes tnpR, blaTEM-
In scientific literature there are examples of antibiotic resistance
1 and aacC5 [10]. These two genes make up a transcription
genes that have been transported to man by bacteria of animal
Table 1 Incompatibility group P plasmids isolated in the Hospital Clínico Universitario, Zaragoza, from 1974 until 1978. Resistance pattern transferred to Escherichia coli K-12: Ap, Tc, Gm, Km, Cm, Sm, Su*
* Ap, Tc, Gm, Km, Cm, Sm, Su: (ampicillin, tetracycline, gentamicin, kanamycin, chloramphenicol, streptomycin, sulfonamide). ‡ Month/year. ¶ Size of all plasmids: ca. 68 kb. ** Cerebrospinal fluid. Table 2 Incompability group M plasmids isolated in the Hospital Clínico Universitario, Zaragoza, from 1976 until 1983. Resistance pattern transferred to Escherichia coli K-12: Ap, Tc, Gm, Tm, Dkb, Nt*
* Ap, Tc, Gm, Tm, Dkb, Nt: (ampicillin, tetracycline, gentamicin, tobramycin, dibekacin, netilmicin).
‡ Month/year. ¶ Size of all plasmids: ca. 73 kb. ** Intensive care units.
origin. It is well known that the correct use of antibiotics in
Other genes have used this system, among them the most
veterinary medicine is of great concern, as it was reported in
prominent are: aadA2, aadB, aacA4, dhfrI, dhfrIIb, dhfrIIc,
the Swann Report (1969) (Report of Joint Comittee on the Use
dhfrV, oxa1, oxa2, pse2, etc. The majority of dhfr genes for
of Antibiotics in Animal Husbandry and Veterinary Medicine,
resistance to trimethoprim are found near integrons that also
Her Majesty’s Stationery Office, London [1969]). Such a
carry the gene sul1, reflecting the joint clinical use of
transfer happened with the use of the aminoglycosides
trimethoprim and sulfamethoxazol. In Tn1696, the close union
apramycin and hygromycin in cattle and pigs respectively. In
of the aadA and aacC1 genes stands out, which implies cross-
fact, through their application, resistant strains emerged, which
resistance on the one hand to streptomycin and spectinomycin
were isolated from animals which had undergone treatment.
and on the other to gentamicin, sisomicin and fortimicin. The
The resistance was plasmid mediated, and genes were detected
stable position of the resistance genes in an integron, present
for apramycin-acetyltransferase [AAC(3)-IV] and hygromycin-
in an R plasmid, assures their dissemination and the capture of
phosphotransferase [APH-(4)-I] enzymes, which inactivated
The mechanism of integron mobility is not well known, but
In 1989 we were able to confirm the isolation of two clinical
there exist at least three possibilities: (i) self mobility, which is
strains (Escherichia coli and Klebsiella pneumoniae) resistant
deduced from the different genetic locations; (ii) by means of
to the abovementioned aminoglycosides, with R plasmids of
transposons of the Tn21 family, in which case they are integrated;
110 kb which carried the aacC4 and aph4 genes. Subsequently,
and (iii), horizontal transfer of integrons by means of R plasmids
four strains of enterobacteria with similar properties were
such as the IncP (pUZ1) and IncW (pSa and R388).
isolated. The analysis of these gene organizations showed thatthey were adjacent and grouped in the same orientation as thosewhich had been isolated from animal origin. The two genes for
resistance to apramycin and hygromycin form an operon and
are associated to IS140 sequences, which implies a transposable
structure [18]. Given that the enzyme AAC(3)-IV inactivatesgentamicin and tobramycin, in addition to apramycin, it
The efficiency of genetic mechanism of transference is reflected
constitutes a potential risk for selecting resistances in clinically
in the spread of multiple resistance to sulfonamides, β-lactam
used aminoglycosides. Furthermore, the six strains isolated,
antibiotics, chloramphenicol, aminoglycosides, etc. The
carrying the plasmids pUZ6734, pUZ6743, pUZ6776, pUZ7852
selection can occur simply through the use of one of the
and pUZ7874, also showed transferable resistance to ampicillin
antimicrobial agents which affects all the genetic determinants.
(β-lactamase TEM-1) and streptomycin [phosphotransferase
An interesting example is the resistance of Haemophilusinfluenzae to chloramphenicol in our geographical region(Aragón) (12%), despite limited clinical use of the drug. Incontrast, 35% were resistant to ampicillin (ApR). And of the
chloramphenicol resistant strains, all except one were also
resistant to ampicillin. This resistance is mediated by
β-lactamases TEM-1. We have not detected any β-lactamase
While trying to characterize the mycobacterial version of the
of type ROB-1, supporting the previously suggested hypothesis
aacC3 gene, the sul3 gene was found, as well as the
of a different ROB-1 reservoir for β-lactam resistance such
determinants int and tnpA. These discoveries allowed us to re-
examine the organization of Tn1696. This 16 kb transposon
chloramphenicol is due to the synthesis of an acetyltransferase
carries the aacC1 gene and the determinants of resistance to
(CAT) which shares biochemical properties with E. coli CAT
sulfamides (sul1), chloramphenicol (cmlA), streptomycin-
II and III. Some of these strains hybridized with a probe for
spectinomycin (aadA1), and mercuric chloride (mer). These
cmlA (a gene present in the integron In4 which is located in
determinants are located in the integron In4, which has a site-
Tn1696), demonstrating permeability changes equivalent to
specific recombination system that permits the capture of
various cassettes of resistance genes [3]. Figure 1 shows the
P. aeruginosa caused by the protein CmlA. In our laboratory,
structure of several integrons, among them In4, including the
we found in H. influenzae the genes which code for the HI1254
cassettes of genes inserted in the variable region [9]. Near
protein, related to the polypeptide CmlA mentioned above, and
the recombination site (res), the int gene was found. It belongs
for the HI1716 protein, which could participate in altering the
to the family of integrases, enzymes essential for recombination.
permeability (Vergara Y, 1996, Ph.D. Thesis, University of
In addition, the cmlA gene, the determinant for non-enzymatic
Zaragoza). Both of these genes have been characterized in the
resistance to chloramphenicol, had a different feature: it has
genome of H. influenzae Rd. On the other hand, there is a low
its own promoter. This does not occur in other integrons of
level of homology between the cmlA gene and the HI1254
protein, which is reflected in weak hybridization with the cmlAFig. 1 Structural features of various naturally occurring integrons. Modified from reference [9]. intI: integrase gene situated in the 5’ conserved segment; sul1: sulfonamide resistance gene situated in the 3’ conserved segment; gene cassettes: aadB, aadA1, etc. with circles (O), which represent recombinaton sites of 59 bp
probe. 37.5% of the H. influenzae strains ApR and CmR were
with enterobacteria, and it seems probable that the species
also resistant to kanamycin, neomycin and lividomycin through
H. parainfluenzae acted as an intermediate.
the production of a phosphotransferase APH(3’)-I. Transfer ofresistance was achieved by electroporation of a competent strain
(E. coli DH5α) with the total DNA of H. influenzae, followed
by selection with ampicillin and kanamycin. Using
electroporation, 180 colonies of E. coli resistant to ampicillinand kanamycin were obtained. DNA was extracted from 9 of
The discovery of strains of Mycobacterium fortuitum producing
the colonies and in each a 7 kb plasmid was found. Restriction
aminoglycoside modifying enzymes [AAC(6’), AAC(3) and
enzimes (EcoRI and PstI) analysis demonstrated that the
APH(3’)] and in particular acetylases subclass AAC(3)-III, has
plasmids were identical. Crude enzyme extracts from the strains
not been related to resistance nor to the presence of plasmids.
showed β-lactamase TEM-1 (isoelectric point, pI = 5.4) activity
A new enzymatic activity that phosphorylated only streptomycin
and 3’-O-phosphotransferase-[APH(3’)-I] activity, which
(APH(6)) was demonstrated in strains of M. fortuitum and M.
modified kanamycin, neomycin and lividomycin [8]. chelonae. Hybridizations with a probe from Streptomyces griseus
The purified phosphotransferase had a molecular weight
(aph-6 gene) were positive not only in the new strains that
of 26,000 Da and a pI = 5.3, with an aminoacid sequence at
produced 6-O-phosphotransferase activity but also in 20 strains
the aminoterminal extreme similar to other phosphotransferases
that did not possess such phosphorylating activity. This suggested
in enterobacteria. Hybridizations with the intragenic probe
that the aph-6 gene was localized on the chromosome, that could
aph(3’)-Ia, demonstrated that the gene in H. influenzae is not
be phenotypically expressed or not, and that did not have any
located in the transposon Tn903, which is a feature that
relation to streptomycin resistance. The investigation was extended
differentiates it from the situation in enterobacteria. In spite of
to other fast growing species (M. smegmatis and M. phlei) and to
this, it was obvious that the determinants of resistance to
slow growing species (M. avium, M. scrofulaceum, M. xenopi
ampicillin, chloramphenicol and kanamycin came mostly from
and M. terrae), verifying in all a positive hybridization, although
E. coli, with peculiarities in genetic reorganization and
to a varying degree. Besides, studying a strain of M. aurum,
phenotypic expression that reflect the change in bacterial host.
the synthesis of an acetylase AAC(2’) with a characteristic profile
In the enterobacteria, the catI, catII and catIII genes are found
was detected. The substrates were gentamicin C1 and C1a,
mainly in IncM plasmids, whereas cmlA and related genes are
netilmicin, 6’-ethyl-netilmicin, tobramycin and kanamycin B,
in IncP plasmids. The coexistance of both types of genes in the
without modifying the 2’-deamino-kanamycins—kanamycin A
genus Haemophilus, situated in plasmids and/or in the
and amikacin—nor the 2’-ethyl-netilmicin (Sánchez-Yangüela
chromosome, shows the heterogeneity of genetic exchange
E, 1990, M.D. Thesis, University of Zaragoza). However, the
genes encoding acetylase AAC(3) subclass III or the related
activation of aac(2’)-Ib in M. smegmatis mc2 155 allowed the
enzyme were not identified, although the enzyme was present in
presence of the gene to be correlated with the resistance to
all the strains of M. fortuitum that were studied and in the majority
lysozyme. Considering this, it is remarkable that the enzyme
AAC(2’)-Ia has been shown to contribute to the acetylation of
In order to detect and characterize the candidate gene aac3,
peptidoglycan (PG) in P. stuartii. Presumably, aminoglycosides
probes derived from genes and transposons coding for antibiotic
are not the true substrate of AAC(2’)-Ia but are acetylated due
resistance were applied to samples of M. fortuitum chromosomal
to their structural similarity with PG [13]. As far as we know,
DNA. A positive hybridization was obtained with a probe
the genes coding for aminoglycoside-modifying enzymes are
containing the aacC1 gene, localized in the previously
not strictly speaking resistance genes in mycobacteria.
mentioned transposon Tn1696. On determining the nucleotide
Paradoxically, if these enzymes are implicated in the
sequence of the hybridizing region, similarities were detected
biosynthesis of the mycobacterial cell wall, they could act as
at the 5’ region of the aacC1 gene. This region comprises the
int gene, which encodes an integrase, a determinant for sitespecific recombination. The mycobacterial sequence alsocontained the sul3 gene, a homologue to the sul1 gene present
in Tn1696 and in other members of the Tn21 family. The region
occupied by the sul3 gene was defined after verifying that itwas flanked by two 880 bp sequences, which were called
The resistance of Streptococcus pneumoniae to penicillin is a
IS6100, and that it encoded a transposase. These mycobacterial
serious problem in Spain. It has become more complicated
insertion sequences contained inverted repetitions of 14 bp,
since the 1980s with an increase in erythromycin resistance,
which differed by only one nucleotide from IS15. The sequence
which rose from 0.9% in 1979 to 10% in 1989. Although
was similar to IS6, and it is closely related to elements isolated
Streptococcus pyogenes remains sensitive to penicillin, in the
from Gram-positive and Gram-negative bacteria. The int and
last few years more and more strains resistant to macrolides
sul3 genes, marked by two IS600 sequences, make up the
have been identified. For this reason, epidemiological
transposon Tn610 [11]. The codons of the IS6100 transposase
gene proved to be similar to those of a gene which encoded the
Of the mechanisms that these streptococci have developed
main antigenic protein of M. tuberculosis. This suggests that
for macrolide resistance, the two fundamental ones are the
this 65,000 Da protein originated in mycobacteria.
production of both methyltransferases, encoded by the erm
The presence of the sul3 gene seemed obvious, since
(A, B, C, TR) genes [2], and the efflux systems, which are the
sulfonamides have been used since 1935 and were applied in
phenotypic expressions of the mefA (S. pyogenes) and mefB genes
the treatment of tuberculosis in the 1950’s. Therefore, finding
(S. pneumoniae), and of the msrA (Staphylococcus epidermidis
the gene in the M. fortuitum FC1 strain suggested a broad
and Staphylococcus aureus) and msrB genes (Staphylococcus
diffusion between this species and others from the genus
xylosus). The methylases encoded by the erm genes cause
Mycobacterium. To test whether this hypothesis was valid,
ribosomal modification at the level of 23S RNA subunit 50S,
hybridizations were carried out with a probe from the int + sul
giving rise to the MLSB phenotype (macrolides, lincosamides,
genes (1.4 kb, plasmid pIPC1::Tn610). All the hybridizations
streptogramin B), either inducible or constitutive. In our medium,
done on 30 strains of M. fortuitum, 21 strains of M. chelonae
the genes detected in Gram-positive cocci belonged to different
and 125 strains of M. tuberculosis were negative. It seemed
classes depending on the genus: Staphylococcus (ermA, ermC,
obvious that we had been lucky in choosing M. fortuitum FC1. ermA + ermC), Enterococcus (ermB) and Streptococcus (ermB),
In fact, initially this strain was selected for its intense acetylating
the expression of which can be inducible or constitutive,
activity AAC(3), and it turned out to house the int, sul3, aph-
irrespective of the class of erm gene [7]. 6 and blaF genes (Timm J, 1994, Ph.D. Thesis, University of
The first description of an efflux mechanism against
Paris VII), and also, as we will describe, the aac-2’-Ib gene.
erythromycin was made in 1989, proving the existence of the MS
A gene encoding an AAC(2’) was cloned from phenotype (14 and 15 membered macrolides, and streptogramine
M. fortuitum. DNA sequencing results identified an
B) as the expression of the msrA gene in S. epidermidis. One year
ORF(aac(2’)-Ib) coding for a putative protein with a predicted
later the gene was characterized and sequenced, demonstrating
molecular mass of 28,400 Da [1]. The deduced AAC(2’)-Ib
that it formed a complex with two other determinants, smp and
protein showed homology to the AAC(2’)-Ia from Providenciastp [16]. In 1992 the msrB gene was cloned and sequenced from
stuartii. The presence of the aac(2’)-Ib in all 34 M. fortuitum
a plasmid from S. xylosus, which expressed the same phenotype
strains was not correlated with any aminoglycoside-resistance
[12]. Using initiators M1 and M2 in one strain of group A
phenotype. This suggested that the product of this gene streptococcus, a 0.4 kb amplification product was obtained, a size
could be implicated in functions other than antibiotic-
compatible with the msrA and msrB genes. A gene which showed
modification, such as the synthesis of intermediate metabolites,
98% homology to the genes characterized in Staphylococcus was
ribosomal proteins, and cell wall components. The insertional
cloned and sequenced from the DNA band (Adrián FJ, 1997,
Ph.D. Thesis, University of Zaragoza).
integron of Tn1696: similarity of the product to transmembrane transport
A new phenotype M has been described in Streptococcus
4. Clancy J, Petitpas JW, Dib-Hajj F, Bergeron J, Yuan W, Cronan M, Kamath
pneumoniae and S. pyogenes, which produces resistance
AV, Retsema JA (1996) Molecular cloning and functional analysis of a
exclusively to 14 and 15 membered macrolides. Investigating
novel macrolide-resistant determinant, mefA, from Streptococcus pyogenes.
this mechanism, a new class of genes was characterized called
mef (for macrolide efflux): mefA in S. pyogenes and mefE in
5. Clancy J, Dib-Hajj F, Petitpas JW, Yuan W (1997) Cloning and
S. pneumoniae [4, 20]. In 1997 a new efflux gene mreA (for
characterization of a novel macrolide efflux gene, mreA, fromStreptococcus agalactiae. Antimicrob Agents Chemother 41:2719–2723
macrolide resistance efflux) was described in Streptococcus
6. Gómez-Lus R, Larrad L, Rubio-Calvo MC, Navarro M, Lasierra MP
(1980) AAC(3) and AAC(6’) enzymes coded by R plasmid isolated in a
In our laboratory, using primers specific for the mefA and
general hospital. In: Mitsuhashi S, Rosival L, Krcmery V (eds) Antibiotic
mefE genes we have confirmed the efflux mechanism in the
resistance. Heidelberg: Springer-Verlag, pp 295–303
majority of M phenotype strains of the species S. pyogenes,
7. Gómez-Lus S, Gómez-Lus R (1993) Detection of new macrolide-
lincosamide-streptogramin resistance phenotypes in gram-positive cocci
S. pneumoniae and different species of the viridans group. The
and its genetic basis. In: Einhorn J, Nord CE, Ragnar Norrby S (eds)
differentiating feature is that the ermB gene is much more
Recent Advances in Chemother. Washington, DC: American Society for
prevalent in the S. pneumoniae population with MLSB
phenotype. Whereas in S. pyogenes the M phenotype expression
8. Gómez-Lus R, Vergara Y (1995) Aminoglycoside resistance in
of the mefA gene is dominant. The M phenotype is infrequent
Haemophilus influenzae. J Chemother 40:97–99
9. Hall RM, Collis CM (1995) Mobile cassettes and integrons: capture and
in pneumococci in our geographical region, in contrast to what
spread of genes by site-specific recombination. Molec Microbiol 15:593–600
has been described in other geographical regions. In almost all
Martín C, Gómez-Lus R, Ortiz JM, García-Lobo JM (1987) Structure
strains the gene mefE was detected.
and mobilization of an ampicillin and gentamicin resistance determinant.
Other resistance mechanisms such as enzymatic
Antimicrob Agents Chemother 31:1266–1270
modification (acetylation, phosphorylation, glycosylation and
11. Martín C, Timm J, Rauzier J, Gómez-Lus R, Davies J, Gicquel B (1990)
Transposition of an antibiotic resistance element in mycobacteria. Nature
nucleotidation) and hydrolysis (esterases), have minor practical
significance in Gram-positive bacteria.
12. Milton ID, Hewitt CL, Harwood CR (1992) Cloning and sequencing of
The transfer of macrolide resistance genes in S. pneumoniae
a plasmid-mediated erythromycin resistance determinant from
is mediated by conjugative chromosomal transposons with a broad
Staphylococcus xylosus. FEMS Microbiol Lett 97:141–148
host spectrum: the genera Streptococcus, Staphylococcus and
13. Payie KG, Rather PN, Clarke AJ (1995) Contribution of gentamicin
2’-N-acetyltransferase to the O acetylation of peptidoglycan in ProvidenciaEnterococcus. Given that resistance to chloramphenicol,
stuartii. J Bacteriol 177:4303–4310
tetracyclines and aminoglycosides (high level) is also transmitted
14. Rivera MJ, Vitoria MA, Navarro M, Robledano L, Chocarro P, Gómez-
by conjugative transposons, these antibiotics can indirectly co-
Lus R (1984) Hospital dissemination among Gram-negative bacillus
select erythromycin, as is the case with Tn1545, which carries the
strains of an IncM plasmid encoding an AAC(3) and a TEM-1
erm, tet, and aph-3’ genes. In contrast, the mutation of the pbp
β-lactamase. Drugs Exptl Clin Res 11:789–795
15. Rivera MJ, Martín C, Gómez-Lus ML, Robledano L, Otal I, Gómez-Lus
genes and transformation by pbp genes from other species,
R (1988) Characterization of a hospital disseminated plasmid encoding
constitutes the well-documented mechanism of penicillin resistance
resistance to gentamicin and other antimicrobial agents. Microbiología
in pneumococci. S. pyogenes exchanges resistance genes by means
of conjugative transposons, for R plasmids and for bacteriophages
16. Ross JI, Eady EA, Cove JH, Cunliffe WJ, Baumberg S, Wooton JC (1990)
(transduction). Nevertheless, studies on resistance gene transfer
Inducible erythromycin resistance in staphylococci is encoded by amember of the ATP-binding transport super-gene family. Molec Microbiol
made in hospitals tend to be retrospective, hence we must try to
examine the process while it is happening, and track the in vivo
17. Rubens CE, McNeill WF, Farrar WE (1979) Transposable plasmid
traffic of R plasmids, transposons and integrons.
deoxyribonucleic acid sequence in Pseudomonas aeruginosa whichmediates resistance to gentamicin and four other antimicrobial agents. JBacteriol 139:877–888
18. Salauze D, Otal I, Gómez-Lus R, Davies J (1990) Aminoglycoside
acetyltransferase 3-IV (aaC4) and hygromycin B 4-I phosphotransferase(hphB) in bacteria isolated from human and animal sources. Antimicrob
Aínsa JA, Martín C, Gicquel B, Gómez-Lus R (1996) Characterization
of the chromosomal aminoglycoside 2’-N-acetyltransferase gene from
19. Smith DI, Gómez-Lus R, Rubio-Calvo MC, Datta N, Jacob A, Hedges
Mycobacterium fortuitum. Antimicrob Agents Chemother 40:2350–2355
RW (1975) Third type of plasmid conferring gentamicin resistance in
Arthur M, Molinas C, Mabilat C, Courvalin P (1990) Detection of
Pseudomonas aeruginosa. Antimicrob Agents Chemother 8:227–230
erythromycin resistance by the polymerase chain reaction using primers
20. Sutcliffe J, Tait-Kamradt A, Wondrack L (1996) Streptococcus
in conserved regions of erm rRNA methylase genes. Antimicrob Agents
pneumoniae and Streptococcus pyogenes resistant to macrolides but
sensitive to clindamycin: a common resistance pattern mediated by an
Bissonnette L, Champetier C, Buisson JP, Roy PH (1991) Characterization
efflux system. Antimicrob Agents Chemother 40:1817–1824
of the non-enzymatic chloramphenicol resistance (cmlA) gene of the In4
Response, activates PLC->IP3,DAG=>Ca++ Similar to M1 mechanism. Sm. Muscles, secretory Agonists Action Bethanechol Stimulates GI and urinary bladder Carbachol + Increases ureteral peristalsis, contraction of detrusor muscle, and decreases Betanechol capacity of urinary bladder (increase in micturation) M3 agonist for salivary gland activation Induces marked salivation and diaphoresis (
Rev. Invest. Mar. 22(1):7-18, 2001 ESTUDIO ELECTROFISIOLÓGICO EN EL CEREBRO DEL CANGREJO MORO ( Menippe mercenaria ) SEMI-INTACTO, NO ANESTESIADO NI RESTRINGIDO . Karina I. Maldonado-León y Oscar H. Hernández Laboratorio de Neurobiología, Centro de Investigaciones en Enfermedades Tropicales. Universidad Autónoma de Campeche. Campeche, México. RESUMEN En este estudio demo
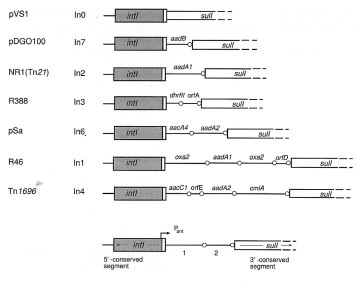 Fig. 1 Structural features of various
Fig. 1 Structural features of various