La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
.p65
Íåéðîíàóêè: òåîðåòè÷í³ òà êë³í³÷í³ àñïåêòè
ÎÐÈòIÍÀËÜͲ²I ÄÎÑ˲²²IÄÆÅÍÍß
BEHAVIORAL DISTURBANCES IN RATS INDUCED VIA THE INTRANIGRAL INJECTION OF
Abstract. According to the experiments carried out on Wistar rats
DSIP is expected to affect behavior and seizure
(250-320 g) with the help of the EEG-, actometry recording
susceptibility. Behavioral consequences of intraSNR
and «open field» indices registration it was shown thatdilateral deltasleep-inducing injection resulted in oligo-
administration are suggested also by known ability of
akinesia, muscle tonus enhancement, decrease of wakefulness
period and deep slow-wave sleep as well as paradoxal on.
homocarnosine in brain cortex, which developed as
Unilateral dilateral deltasleep-inducing administration(5-20 nmole) into substantia nigre reticulation resulted in the
a result of activation of GABA-decarboxylase and
appearance of dose-dependent contralateral circlings, which
decrease of GABA-transaminase activity [24, 25].
were blocked by naloxone (1.0 mg/kg). Haloperidol
Hence the aim of the present investigation was to
(1.0 mg/kg) caused the potentiation on keeping anuncomfortable position in animals with bilateral intra
observe behavioral disorders induced by intraSNR
substantia nigre reticulation dilateral deltasleep-inducing
DSIP, with the emphasis on PD-connected deteriorations
administration, while naloxone (1.0 mg/kg) markedly reduced
and DSIP- induced effects, such as locomotion, sleep-
this index. Analogous dilateral deltasleep-inducing intrastriatalinjection failed to induce the same behavioral disturbances.
wakefulness cycle, and antiseizure activity.
Intranigral dilateral deltasleep-inducing administration was
followed by decreasing of seizure susceptibility to intra
The experiments were carried out on male Wistar
picrotoxin administration (2,0 mg/kg).
rats with the weight of 250-320 g. All the animals were
Key words: delta sleep-inducing peptide, substantia nigra reticulata,
kept at a constant room temperature of 22oC with
circlings, naloxone, haloperidol, picrotoxin, seizures
12 hr artificial dark/light cycle and free access to astandard diet and tap water.
Substantia nigra (SN) continues to attract attention
Procedures involving animals and their care were
as a structure with the lowest threshold of a certain
conducted according to University guidelines that
reaction, such as rotation, which is considered to be
comply with international laws and policies [European
a “hemiparcinsonian” state [17]. Bilateral reduction
Community Council Directive 86/609, OJ L 358, I,
of SN dopamine output underlies behavioral
December 12, 1987; National Institute of Health
deteriorations, which are regarded as an experimental
Guide for Care and Use of Laboratory Animals, US
equivalent of Parkinsonian disease (PD) [27].
Inhibition of the dopaminergic neurons activity of SN
Stainless steel guide cannulas (external diameter
pars compacta might be achieved via pharmacological
0.5 mm) were implanted into the SNR and striatum
modulation of SN reticular (SNR) neurons [7]. Thus,
under hexenal anesthesia (100 mg/kg) in accordance
bilateral stimulation of GABA-A or GABA-B receptors
with the rat brain atlas [29]: (AP=-4.0; L=2.5; H=8.0
with muscimol and baclofen in SNR counteracted the
and AP=0.8; L=3.0; H=4.5, correspondently). The
evoked dopamine release [2]. Besides, baclofen
cannulas were implanted bilaterally so that they did
reduced, while muscimol intensified locomotor activity
not cross the upper boundary of the structure under
It is interesting to note that SN is a structure, which
In order to identify the various stages of sleep
converts peptidergic inputs into neuromediator outputs
recording electrodes were implanted into the
[10, 19, 20]. Hereby, peptidergic mechanisms are
hippocampus (AP=-4.0; L=2.5; H=3.5), sensorimotor
supposed to be also involved in the PD development,
cortex and the neck muscles; an indifferent electrode
and opiate- induced muscle rigidity is the most
was attached to the nasal bones. Quick-soliding dental
intensively discussed in that respect [18, 26].
plastic material was used for the fixation of the cannulas
Several compounds, including those ones of peptide
and electrodes to the skull. After the operations the rats
nature, applied intranigrally caused antiseizure effects
were caged in groups of 5-10 animals. Since a week after
[5, 7, 9,-11, 23]. To some extent the intraSNR test could
the surgery, the rats had been handled daily and adapted
be regarded as discriminate for antiepileptics [8, 38]. In
our experiments intranigral administration was used as
Investigations were then started 1-2 weeks after the
a screening test for antiepileptic potency of structural
surgery. All the observations were performed from
analogues of delta sleep-inducing peptide (DSIP) [33].
11.00 am to 5.00 pm. The number of rats in each
The recent investigations revealed the DSIP
group was 5-12. The analysis of circlings was confined
ability to block seizures induced via stimulations of
to estimate the number of rotations. It was performed
the receptors of excitatory aminoacids [35, 37]. In this
during the period of 20 min following the DSIP
respect it is of interest to note that stimulation of
injection into the left SNR. Locomotive activity was
intraSNR excitatory aminoacid receptors resulted in
evaluated quantitatively by means of seismorecorded
generation of seizure activity [33]. Hence, intraSNR
data registration for a period of 2 minutes after the
Êîðåñïîíäåíö³ÿ: Å.Â. Êîáîëºâ, Ëüâ³âñüêèé íàö. ìåä. óí³âåð.,âóë. Ïåêàðñüêà, 69, Ëüâ³â, 79010, Óêðà¿íà
Table 1. Effects of deltasleep-inducing injection (DSIP) and its combination with naloxone and haloperidol
(*) - P<0.05 when compared with DSIP 5 nmol group (ANOVA test)
placement of an animal on a movable floor. In
USA) were injected intra in a dose of 1.0 mg/kg,
addition «open field» data was determined by calculating
10 min before the DSIP administration.
the number of squares crossed during a period of
At the end of the experiments, the rats were
2 minutes after placing the animal in the center of the
anesthetized with pentobarbital sodium and perfused
area3. Catalepsy was evaluated by determining the
with paraformaldehyde. Frozen sections (32 mm) of
duration of retention of an uncomfortable posture
the brain were then prepared and every alternate
[28]. Muscle tonus was estimated by clinically testing
section mounted on gelatin-coated slides, stained with
and recording the resistance of muscles to passive limb
neutral red, covered with cover-slip, and examined by
adduction. The number of animals with stereotype
light microscopy. In all rats used in the analysis of
behavior (i.e. sniffing, licking, and gnawing) was
the data the cannula traces and electrodes were
noted as an informative index for the identification
identified at the appropriate location.
For data analysis, when appropriate, parametric
The investigations of the sleep-wakefulness cycle
statistic (Student’s t-test, analyses of variance) or the
non-parametric statistic (Wilcoxon U-test) were
electrographical and behavioral indices recorded after
used. Krusckall-Wallis test was used in case of seizure
the animals had been placed inside a sound-isolated
severity estimation. P values <0.05 were considered
box, with a constant level of artificial illumination.
Actometry and EEG-recordings were then carried out
via signals sampling at 256 samples/s with the use of
DSIP administration into the left SNR caused the
a data acquisition board (National Instruments,
manifestations of contralateral circlings, the intensity
USA), and stored for off-line analysis. The signals
of which depended on the dose peptide used. Circlings
were band pass filtered, only frequencies between
0.5-40 Hz were allowed to pass. The polygraph
administration, and their maximal intensity was
records were inspected visually and epochs containing
observed during 2-4 min after the moment of their
artifacts discarded. Actometry recording was performed
appearance. Circlings disappeared in 10-13 min after
during the sleep-wakefulness cycle investigation.
the beginning of the observation (tab. 1). Periodical
EEG was evaluated minimally during 50 sec [30]. Thestandard duration of the separate phases was ultimatelydetermined manually. Two phases of slow sleep were
Tabl. 2. Effects of deltasleep-inducing injection
determined in the followig way1: the light phase was
(DSIP) and its combination with naloxone and
characterized by the appearance of unstable, relatively
haloperidol on the duration of uncomfortable position
low-amplitude activity with separate theta and delta
waves, the occurrence of which did not exceed 180
mcV; single alpha-rhythm spindles were also noted
at this stage. The second phase - the deep slow wave
stage was characterized by an increase in the number
and amplitude of theta and delta waves: up to 200 mcV.
The latency in both falling asleep and paradoxal sleep
Microinjections of DSIP (synthesized and purified
by the method described above [33]) were conducted
under mild ether anesthesia after fixing the animals
in a stereotaxic device. Peptide was dissolved in saline
in a volume of 1.0 mcl and injected in dosages of 5.0;10.0 and 20.0 nmole. The speed of injections was
0.5 mcl/min. Analogous administration of saline was
performed on rats in the control groups. Haloperidol
(«Gedeon Richter», Hungary) and naloxone («Sigma»,
P was calculated using the ANOVA test.
Fig. 1. The influence of deltasleep-inducing injection injection into substantia nigre reticulation and caudate nuclei on locomotor
A: Ordinate: the locomotor activity related pertaining to the indices of horizontal and vertical activity in animals treated with saline
(control, 100%); Abscissa: time after pharmacons administration (hours); B: Ordinate: the same as in «A»; Abscissa: I - DSIP intraSNR administration in dose of 5,0 nmol; II- and III- DSIP administration into rostral part of n.caudatus in dosage of 5,0 and 10,0nmol correspondently.
The number of experimental animals are presented inside the bars. #-P<0,05 and ##-P<0,01 in comparison with the control group.
sniffings were observed in 2 of 8 animals and 3 animals
motor functions, restricted limb movements and
demonstrated chewing during their circlings. These
decrease in size of steps taken were registered.
reactions did not differ from those in the control group,
Significant resistance in passive limb adduction and
where stereotype sniffings were recorded in 3 of 7 rats.
increase in muscle tonus occurred. Ptosis, periodical
After the placement of the rats in an uncomfortable
gnawing and sniffings were registered in 3 of 8
position (on the side or back) they retained it for
animals. The mentioned above behavioral disturbances
17.9+3.2 sec a significantly longer period in comparison
were observed during a period of 60-80 min after the
with the analogous data obtained from the animals in
DSIP microinjections with a continual decrease in the
the control group (1.4+0.2 sec; F(1,12)=26.46, p<0.001;
indices investigated later on. The absence of differences
(tab. 2). After the preliminary naloxone administration
between the experimental and control groups as well
the animals failed to keep the uncomfortable posture
as with the initial level was observed 3.5-6.0 hrs from
imposed to them (F(1,12)=0.03, p>0.05; Table 2) and
the moment of DSIP intra-SNR administration
no circling behavior was observed (tab. 1). Stereotype
(fig.1,A). An intranigral DSIP administration in a
sniffings and gnawings were displayed by 3 of 7 rats.
dose of 5 nmole as well as a DSIP injection into the
The DSIP injection which was administered after a
rostral parts of the nucleus caudatus in a dose of 10
preliminary dose of haloperidol, resulted in much
nmole did not result in a significant decrease of
longer retention of uncomfortable posture - 179+35
locomotive activity in the animals (fig.1,B) and did
sec (F(1,12)=25.74, p<0.001; Table 2). When the
animals eventually regained a vertical position, markedly
The sleep-wakefulness cycle investigations carried
reduced locomotive activity was recorded - the rats
out on 5 rats with intranigral DSIP microinjections
remained immobile during all the period of observation
revealed a significant decrease in active wakefulness
(tab. 1). Paroxysmal gnawings were observed in 2 of
(F(1,18)=36.13, p<0.001) and prolonged deep slow
8 rats and 2 rats displayed stereotype sniffings.
wave sleep and paradoxal one (F(1,18)=11.08, p<0.01
3-10 min after the DSIP injection into the SNR
and F(1,18)=8.00, p<0.05), correspondently, compared
paroxysmal arrest reactions with a duration of 10-20
to the analogous indices of the animals in the control
sec to 5.0-6.5 min were observed. After the placement
group (injected intranigrally with saline) (fig. 2,A).
of the rats into the center of the «open field», 4 of 10
These changes were recorded in the background of the
animals remained as placed throughout observation
decrease in locomotive activity (fig. 2,B).
(2 min). The average number of squares crossed
Picrotoxin (2,0 mg/kg) produced the first convulsive
decreased significantly in comparison with the
reactions at average 22,1+ 1,0 min after dosage in
analogous index in the animals of the control group
control animals (saline injected into SNR). The
which decreased respectively from 547+130 to 129+46;
intensity of seizures increased over the next
F(1,18)=9.01, p<0.01; (fig.1,A) as well as with its
10-15 min, and generalized convulsive attacks occurred
initial value in the animals of the experimental group.
in 15 of 17 rats with latent period of 35,0+ 1,5 min.
Furthermore, a significant decrease in vertical bars
The mean intensity of convulsions was 4,0+0,1
was observed (fig.1,A) compared both with the
points. The other two rats had clonic convulsions of
background level and with the activity in the control
group (from 27.3+2.5 to 10.9+4.5 (F(1,18)=10.15,
Convulsant treatment of rats given bilaterally
p<0.01). During locomotion the slowing-down of
intranigrally DSIP led to the formation of seizures
Fig. 2. Sleep-wakefulness cycle characteristics under condi-
Fig.3. Seizure manifestations induced by picrotoxin (2,0
tions of deltasleep-inducing injection injection into substantia
mg/kg. i.p.) administration to rats treated intranigrally substantia
nigre reticulation with deltasleep-inducing injection (10 nmol).
A: Ordinate: seconds; Absciss: 1- wakefulness; 2- latency of
Ordinate- indices under investigation in % pertained to
sleep precipitation; 3- superficial, and 4- deep stages of slow-
corresponded ones in the control group of animals (100 %).
wave sleep; 5- latency of paradoxal sleep precipitation;
#-P<0.05; ###-P<0,001 in comparison with the control
6- paradoxal sleep; B: Ordinate: actometry data; Abscissa: time
after pharmacons administration (minutes).
#-P<0,05 in comparison with the control group.
with a latent period which was 24% greater than that
excluded. The opioid nature of other DSIP-induced
in control animals (F(1,25)=6.61, p<0.05) (fig. 3).
effects, namely sleep induction, were also suggested [1].
Rats showed intense clonic convulsions of the
In the case of intranigral DSIP injection a
muscles of the trunk and hindlimbs; two of ten
significant decrease in wakefulness and increase in
animals developed generalized convulsive seizures.
both deep slow-wave and paradoxical sleep occurred.
The latency of generalized clonic-tonic fits exceeded
This data testifies to the fact that SNR structures do
that one in control group by 61,0% (F(1,25)=62.80,
take part in deep slow-wave and paradoxal sleep,
p<0.001). The mean severity of seizures was 20%
which are known to enlarge after systemic DSIP
lower than that in control animals (H=14.52, p<0,001).
administration [22, 39]. It should be noted that the
Our investigations have shown that unilateral DSIP
muscle tonus reduction, which is characteristic of
administration into the SNR caused contralateral
precipitation of the paradoxal sleep [16] is apparently
rotations of a dose-dependent character. The efficacy
contrary to the muscle rigidity induced after DSIP
of naloxone in relatively low dosage (1.0 mg/kg) in
intra-SNR administration. Such a contradiction
preventing any circlings suggests that the opioid
could be explained by the periodical DSIP-induced
mechanisms activated by the intranigral DSIP testify
activation of the endogenous system, which controls
to their development. Haloperidol was also effective in
the decreasing of muscle tonus (presumably - part
this way. But the simultaneous participation of
of nuclei involved in PS-precipitation). Such an
dopaminergic mechanisms as alternative cause of the
induction could be secondary resulting from a
circlings could hardly be suggested because the
consequence of the primary mechanisms of muscle
haloperidol and DSIP-treated animals retained their
uncomfortable posture significantly longer than those
The activation of the mechanisms of paradoxal
treated with separately administrated drugs. It might be
sleep is shown to cause anticonvulsive effects [3, 36].
that the locomotive disabilities were influenced by the
According to it [16] the turn on-off mechanisms of
circlings. Therefore, the potentiation of haloperidol-
paradoxical sleep are determined by noradrenergic
induced catalepsy by intra-SNR DSIP could be
brain systems. On the other hand, it has been shown
supposed and the neuroleptic mode of DSIP-induced
that the antiepileptic effects of intranigral muscimol
effects could be suggested. This suggestion is supported
are blocked by yohimbine (alpha-2-adrenoreceptor
by the data showed a marked increase of DSIP-like
antagonist), which results in the disinhibition of
immunoreactivity in the hypothalamic region after
noradrenergic terminals [6, 31]. Thus, it could be
systemic haloperidol administrations [40].
assumed that the antiseizure effect of intranigral DSIP
It should be noted that stereotypic sniffings and
administration, together with the application of other
gnawings were observed after the haloperidol application
drugs, may be achieved via the paradoxal sleep
following the intranigral opioid peptides administration
mechanisms disinhibition as a result of the presynaptic
[18, 26]. Therefore, the participation of opioid
inhibition of corresponding noradrenergic
mechanisms in such behavioral changes is not to be
mechanisms. From the viewpoint of the above
discussion the ability of DSIP to modulate the
ðàëüíîå ââåäåíèå äåëüòàñîí-èíäóöèðóþùåãî ïåï-
adrenoreceptor activity [14] as well as dopaminergic
òèäà òàêæå óìåíüøàëî ñóäîðîæíóþ ãîòîâíîñòü
êðûñ â îòíîøåíèè âíóòðèáðþøèííîãî ïðèìåíå-
íèÿ ïèêðîòîêñèíà (2,0 ìã/êã). (Íåéðîíàóêè: òåîð.
Hence, the participation of the opiatergic and
êëèí. àñï.— 2006. — Ò. 2, ¹ 1-2. — Ñ. 14-19).
catecholaminergic mechanisms in the creation of
Êëþ÷åâûå ñëîâà: äåëüòàñîí-èíäóöèðóþùèé ïåï-
behavioral effects of intra-SNR administered DSIP
òèä, ðåòèêóëÿðíàÿ ÷àñòü ÷åðíîãî âåùåñòâà, ðîòà-
was supposed. Besides, DSIP intensifies GABA synthesis
öèè, íàëîêñîí, ãàëîïåðèäîë, ïèêðîòîêñèí, ñóäî-
[24, 25], which could be suggested as the motive of the
display of the stereotype behavior described above [6]. The role of DSIP-induced restriction of elaboration of
ILs can not be excluded [41]. Finally, the interaction
Ïîâåä³íêîâ³ ïîðóøåííÿ, âèêëèêàí³ ó ùóð³â
between opioid, GABA systems realized with thealpha-2-adrenoreceptors was reported [4, 19].
âíóòð³øíüîí³ãðàëüíèì çàñòîñóâàííÿì
The above- mentioned scheme of consequences is
äåëüòàñîí-³íäóêóþ÷îãî ïåïòèäó
congruent with the antiepileptic mechanisms realized
Ó äîñë³äàõ íà ùóðàõ ë³í³¿ ³ñòàð (250- 320 ã) çà
via action of different substances upon SNR. As, for
äîïîìîãîþ ìåòîä³â ÅÅÃ-ðåºñòðàö³¿, àêòîìåòð³¿,
the cause of antiepileptic effects such mechanisms as
äîñë³äæåíü ïîêàçíèê³â ó «â³äêðèòîìó ïîë³» áóëî
activation of GABA mediation [6, 17, 31], kappa-
âñòàíîâëåíî, ùî á³ëàòåðàëüíå çàñòîñóâàííÿ äåëü-
opioid receptors [5] along with blocking of excitatory
òàñîí-³íäóêóþ÷îãî ïåïòèäó âèêëèêàëî, îë³ãî- àê-
mechanisms [9], substance P [11] should be mentioned.
³íåç³þ, çá³ëüøóâàëî òîíóñ ìÿç³â, çìåíøóâàëî ïå-
Besides, effects of different peptides intranigrally is
ð³îä íåñïàííÿ ³ âèêëèêàëî ãëèáîêèé ïîâ³ëüíîõâè-
ëüîâèé ñîí, à òàêîæ ïàðàäîêñàëüíèé ñîí. Îäíî-
resulted in antiseizure action; kyotorphin and its
ñòîðîííº çàñòîñóâàííÿ äåëüòàñîí-³íäóêóþ÷îãî
structural analogues [13], somatostatin, neurotensin
ïåïòèäó (5-20 íìîëü) â ðåòèêóëÿðíó ÷àñòèíó ÷îð-
[34] as well as activation of benzodiazepine receptors
íî¿ ðå÷îâèíè âèêëèêàëî ïîÿâó êîíòðëàòåðàëüíèõ
[23] heve been described. Mentioned induction of
ðîòàö³é, ÿê³ áóëè äîçà-çàëåæíèìè òà áëîêóâàëèñü
neurotransmitter-involved mechanisms is attracted
çà äîïîìîãîþ íàëîêñîíó â äîç³ 1,0 ìã/êã. Ãàëîïå-
for the explanation of the action of each of peptides.
ð³äîë (1,0 ìã/êã) ïîòåíö³þâàâ çäàòí³ñòü âíóòð³ø-
íüîí³ãðàëüíîãî çàñòîñóâàííÿ äåëüòàñîí-³íäóêó-
It should be stressed that neurodegeneration,
þ÷îãî ïåïòèäó âèêëèêàòè òðèâàëå óòðèìóâàííÿ
which is pertinent to PD, is opposite to effect of
òâàðèí â íåçðó÷í³é ïîç³, â òîé ÷àñ ÿê íàëîêñîí
neuroprotection induced by DSIP [37]. Hence, to
çìåíøóâàâ âêàçàíèé åôåêò äåëüòà-ñîí ³íäóêóþ÷î-
support the role of DSIP in PD related disturbances
ãî ïåïòèäó. Àíàëîã³÷íå âíóòð³øíüîñòð³àðíå çàñ-
pathogenesis means to support the secondarily character
òîñóâàííÿ äåëüòàñîí-³íäóêóþ÷îãî ïåïòèäó íå âèê-
of DSIP-like neuropeptides rising in zone of primarily
ëèêàëî ïîä³áíèõ ïîâåä³íêîâèõ ïîðóøåíü. Âíóòð³-
øíüîí³ãðàëüíå ââåäåííÿ äåëüòàñîí-³íäóêóþ÷îãî
increasing of neurotoxicity. In this case all DSIP-
ïåïòèäó òàêîæ çìåíøóâàëà ñóäîìíó ãîòîâí³ñòü
ùóð³â ùîäî âíóòð³øíüîî÷åðåâèííîãî çàñòîñó-
hypercompensative state of peptidergic systems in
âàííÿ ï³êðîòîêñèíó (2,0 ìã/êã). (Íåéðîíàóêè: òåîð.
SNR. In the course of exhausting of their activity,
êë³í. àñï.— 2006. — Ò. 2, ¹ 1-2. — Ñ. 14-19).
neurotoxicity resulted in stable degenerative neuronal
Êëþ÷îâ³ ñëîâà: äåëüòà ñîí-³íäóêóþ÷èé ïåïòèä,
deteriorations and substitution of «compensative»
ðåòèêóëÿðíà ÷àñòèíà ÷îðíî¿ ðå÷îâèíè, ðîòàö³¿,
íàëîêñîí, ãàëîïåð³äîë, ï³êðîòîêñèí, ñóäîìè
symptoms by stable «pathogenic» ones.
1. Akahiro N., Masaya N., Toko S. et al.//Eur. J.Pharmacol.
Ïîâåäåí÷åñêèå íàðóøåíèÿ, âûçâàííûå ó
— 1988. — Vol. 155. — P. 240-253.
êðûñ âíóòðèíèãðàëüíûì ïðèìåíåíèåì
2. Balon N., Kriem B., Weiss M. et al. // Brain Res. — 2002.
äåëüòàñîí- èíäóöèðóþùåãî ïåïòèäà
3. Boldy-Moulnier M. Inter-relationships between sleep and
epilepsy. Recent Advances in Epilepsy. Number three.
 îïûòàõ íà êðûñàõ ëèíèè Âèñòàð (250- 320 ã) ñ
Eds.Pedley T.A., Meldrum BS. ôìùò avon (G.B.): Bath
ïîìîùüþ ìåòîäîâ ÝÝÃ-ðåãèñòðàöèè, àêòîìåòðèè,
èññëåäîâàíèÿ ïîêàçàòåëåé â òåñòå «îòêðûòîãî ïîëÿ»
4. Bernasconi R., Aryces D., Martin P. et al. //Naunyn-Schmiedebergs
áûëî óñòàíîâëåíî, ÷òî áèëàòåðåëüíîå ïðèìåíå-
Arch. Pharmacol. — 1986. — Vol. 334, Suppl. — P. 47.
íèå äåëüòàñîí-èíäóöèðóþùåãî ïåïòèäà âûçûâàëî
5. Bonhaus D.W., Rigsbee L.S., McNamara J.O. //Brain Res.
— 1987. — Vol. 405. — P. 358-363.
îëèãî-àêèíåçèþ, óâåëè÷èâàëî ìûøå÷íûé òîíóñ,
6. Bonhaus D.W., McNamara J.O. Brain Res. — 1988. — Vol.
óìåíøàëî ïåðèîä áîäðñòâîâàíèÿ è âûçûâàëî ãëó-
áîêèé ìåäëåííîâîëíîâîé ñîí, à òàêæå ïàðàäîê-
7. Celada P., Paladini C.A., Tepper J.M. // Neuroscience. —
ñàëüíûé ñîí. Îäíîñòîðîííåå ââåäåíèå äåëüòàñîí-
èíäóöèðóþùåãî ïåïòèäà (5-20 íìîëü) â ðåòèêó-
8. Chen L.S., Millington D.S., Maltbay D.A., McNamara
ëÿðíóþ ÷àñòü ÷åðíîãî âåùåñòâà âûçûâàëî ïîÿâëå-
J.O.// Neuropharmacology. — 1989. — Vol. 28. —
íèå êîíòðëàòåðàëüíûõ ðîòàöèé, êîòîðûå áûëè
äîçà-çàâèñèìûìè è áëîêèðîâàëèñü ñ ïîìîùüþ
9. De Sarro G.O., Meldrum B.C., Reavill C. // Eur. J.
Pharmacol. — 1986. — Vol. 106. — P. 175-179.
íàëîêñîíà â äîçå 1,0 ìã/êã. Ãàëîïåðèäîë (1,0 ìã/êã)
10. Garant D.S., Iadorola M.J., Gale K. //Soc. Neurosci.
ïîòåíöèðîâàë ñïîñîáíîñòü âíóòðèíèãðàëüíîãî
Abstr. — 1982. — Vol. 8. — P. 281-282.
ïðèìåíåíèÿ äåëüòàñîí-èíäóöèðóþùåãî ïåïòèäà
11. Garant D.S., Iadorola M.J., Gale K. // Brain Res. — 1986.
âûçûâàòü äëèòåëüíîå óäåðæàíèå êðûñ â íåóäîáíîé
äîçå, â òî âðåìÿ êàê íàëîêñîí óìåíüøàë óêàçàí-
12. Gershtein L.M., Dovedova E.L.//Neurochem. Res. — 1999.
íûé ýôôåêò äåëüòàñîí-èíäóöèðóþùåãî ïåïòèäà.
Àíàëîãè÷íîå âíóòðèñòðèàðíîå ïðèìåíåíèå äåëü-
13. Godlevsky L.S., Shandra A.A., Mikhaleva I.I. et al.// Brain
òàñîí-èíäóöèðóþùåãî ïåïòèäà íå âûçûâàëî ïî-
Res. Bull. — 1995. — Vol. 37. — P. 223- 226.
14. Graf M.V., Schoenenberger G.A.//Biol. Chem. Hoppe-
äîáíûõ ïîâåäåí÷åñêèõ íàðóøåíèé. Âíóòðèíèã-
Seyler. — 1985. — Vol. 366. — P. 795.
15. Henderson J.M., Watson S.H. // Brain Res. Bull. — 2003. —
28. Ossowska K., Wedzony K., Wolfarth G.//Pharm. Biochem.
Behav. — 1984. — Vol. 21. — P. 825-831.
16. Hobson J.A., McCarley R.W., Wyzinski P.W. //Science. —
29. Paxinos G., Watson C. The Rat Brain in Stereotaxic
Coordinates. Academic Press Inc., Sydney, 1998.
17. Iadorola M.J., Gale K. // Science. — 1982. — Vol.218. —
30. Pellegro T., Monti J.M., Baglietto J. et al.// Sleep. — 1984. —
18. Iwamoto E.T., Way E.L. //J.Pharmacol. Exp. Ther. —
31. Platt K., Butler L.S., Bonhaus D.W. et al.//J.Pharmacol.
1977. — Vol. 203. — P. 347- 359.
Exp. Ther. — 1987. — Vol. 241. — P. 751-754.
19. Kalivas P.W.// Neurosci. and Behav. Reviews. — 1985. —
32. Prudchenko I.A., Stashevskaya L.V., Mikhaleva I.I. et al.// Russian
J. of Bioorganic Chemistry. — 1993. — Vol.19. — P.23-32.
20. Kamata K.// Japan J.Pharmacol. — 1987. — Vol. 45. — P.
33. Sawamura A., Hashizume K., Yoshida K., Tanaka T.//
Brain Res. — 2001. — Vol. 911. — P. 89-95.
21. Karmanova I.G., Maximuk I.P., Voronov I.B. et al.//
34. Shandra A.A., Godlevsky L.S., Vastyanov R.S., Panenko A.V./
J.Evolutionary Biochem. and Physiol. (Moscow, in Russian). —
/ Physiol.J. (Kiev, Ukraine). — 1993. — Vol.39. — P.76-81.
35. Shandra A.A., Godlevskii L.S., Brusentsov A.I. et al.//
22. Kimura M., Inove S.//Psychopharmacology. — 1989. —
Neurosci. Behav. Physiol. — 1998. — Vol. 28. — P. 521-526
36. Shouse M.N., Staba R.J., Saquib S.F. et al.//Brain Res. —
23. King P.H., Shin Ch., Mansbach H.H. et al.//Brain Res. —
37. Stanojlovic O., Zivanovic D., Mirkovic S. et al.//Pharmacol.
24. Mendzheritsky A.M., Mackletsova M.G., Karouchina I.M.//
Biochem. Behav. — 2004. — Vol. 77. — P. 227-234.
Neurochemistry (FSU, in Russian). — 1987. — Vol. 6. — P.
38. Turski L., Andreus G.S., Loschmann P.A. //Brain Res. —
25. Mendzheritskii A.M., Lysenko A.B., Uskova N.I. et al.//
39. Ursin R., Larsen M.// Neurosci. Lett. — 1983. — Vol. 40. —
Neurosci. Behav. Physiol. — 1997. — Vol.27. — P. 714-717.
26 Morelli M., Dichiara G.// Brain Res. — 1985. — Vol.341. —
40. Wahlestedt C., Ekman R., Heilig M. et al.//Eur. J.
Pharmacol. — 1989. — Vol. 159. — P. 285-289.
27. Obeso J.A., Rodriguez-Oroz M.C., Rodriguea M. et al.//
41. Yehuda S., Mostofsky D.//Peptides. — 1993. — Vol. 14. —
Trends Neurosci. — 2000. — Vol. 23(Suppl.), — P.8-19.
Íàä³éøëà äî ðåäàêö³¿: 07.05.2006
AMERICAN GINSENG: THE ROOT OF NORTH AMERICA'S MEDICINAL HERB TRADE A TRAFFIC North America report May 1998 Ginseng ( Panax spp.) is arguably the most revered medicinal plant in traditional Chinese medicine and is quickly becoming one of the most popular herbs in Western markets. In the United States, where the market for medicinal botanicals is US$3 billion (CA$4.3 billion) a
Lymphoma is considered to be the most chemo-responsive cancer in cats and treatment with multi-agent chemotherapy is associated with the longest survival times. Common protocols include ACOPA and Madison Wisconsin, but both utilize the same chemotherapy agents. The induction part of the treatment protocol ranges from 21-25 weeks. The goal of induction chemotherapy is to induce a remission whi
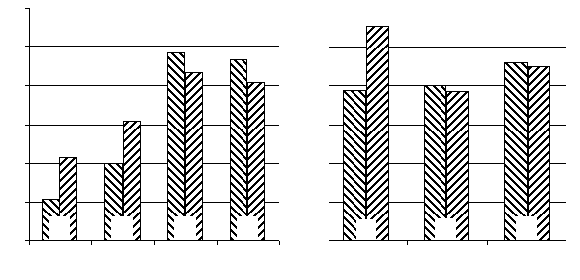 Fig. 1. The influence of deltasleep-inducing injection injection into substantia nigre reticulation and caudate nuclei on locomotor
A: Ordinate: the locomotor activity related pertaining to the indices of horizontal and vertical activity in animals treated with saline
(control, 100%); Abscissa: time after pharmacons administration (hours); B: Ordinate: the same as in «A»; Abscissa: I - DSIP intraSNR administration in dose of 5,0 nmol; II- and III- DSIP administration into rostral part of n.caudatus in dosage of 5,0 and 10,0nmol correspondently.
Fig. 1. The influence of deltasleep-inducing injection injection into substantia nigre reticulation and caudate nuclei on locomotor
A: Ordinate: the locomotor activity related pertaining to the indices of horizontal and vertical activity in animals treated with saline
(control, 100%); Abscissa: time after pharmacons administration (hours); B: Ordinate: the same as in «A»; Abscissa: I - DSIP intraSNR administration in dose of 5,0 nmol; II- and III- DSIP administration into rostral part of n.caudatus in dosage of 5,0 and 10,0nmol correspondently.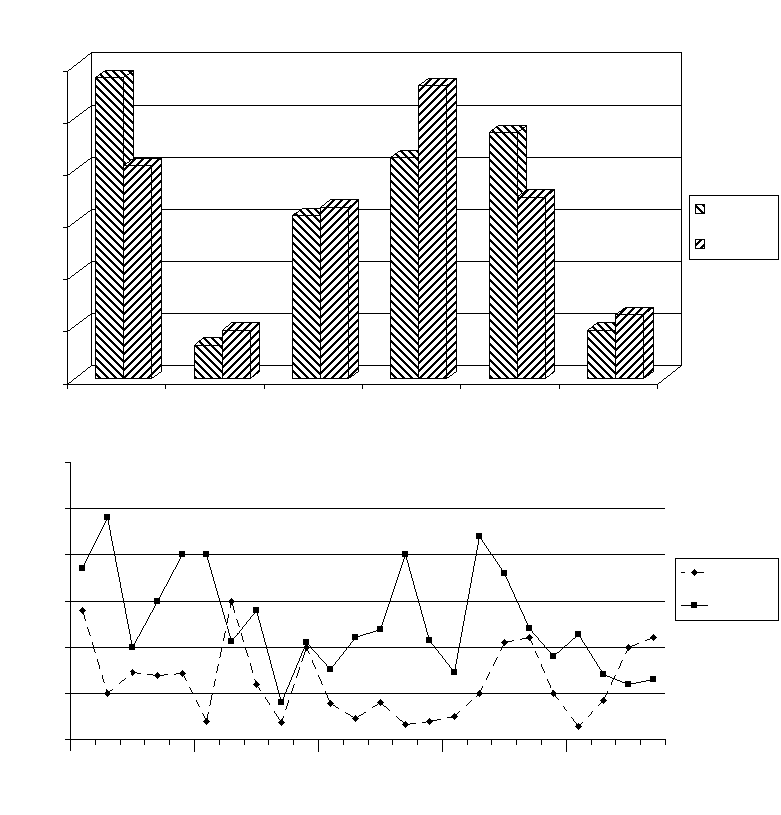
 Fig. 2. Sleep-wakefulness cycle characteristics under condi-
Fig.3. Seizure manifestations induced by picrotoxin (2,0
tions of deltasleep-inducing injection injection into substantia
mg/kg. i.p.) administration to rats treated intranigrally substantia
nigre reticulation with deltasleep-inducing injection (10 nmol).
Fig. 2. Sleep-wakefulness cycle characteristics under condi-
Fig.3. Seizure manifestations induced by picrotoxin (2,0
tions of deltasleep-inducing injection injection into substantia
mg/kg. i.p.) administration to rats treated intranigrally substantia
nigre reticulation with deltasleep-inducing injection (10 nmol).