La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Micoleao.org.br
International Journal of Primatology [ijop]
International Journal of Primatology, Vol. 24, No. 6, December 2003 ( C 2003)Endocrine Monitoring of Wild Dominant and Subordinate Female Leontopithecus rosalia Jeffrey A. French,1,5 Karen L. Bales,2,4 Andrew J. Baker,3 and James M. Dietz2 Received February 6, 2003; revision May 22, 2003; accepted June 2, 2003In captive callitrichid primates, female reproductive function tends to varywith social status. However, little is known about the interplay between thesefactors in wild groups. We report observations on normative ovarian functionin dominant and subordinate female golden lion tamarins (Leontopithecusrosalia) living in wild groups. We monitored ovarian status by measuring, viaenzyme immunoassay, concentrations of excreted pregnanediol glucuronide(PdG) and estrone conjugates (E1C) in fecal samples collected noninvasivelyfrom individuals in social groups in the Poc¸o das Antas Biological Reserve,Rio de Janeiro State, Brazil. Dominant breeding females demonstrated steroidlevels similar to those previously reported for wild cotton-top tamarin females,with statistically significant rises during pregnancy. The duration of elevationof fecal steroids in breeding females was ca. 4 mo, which corresponds withestimates of gestation from captive studies. Low steroid concentrations fromDecember to June suggest a seasonally-related period of infertility in femalegolden lion tamarins. Dominant and subordinate females demonstrated sev-eral differences in endocrine function. In general, younger females living inintact natal family groups showed no evidence of ovarian cyclicity. We notedendocrine profiles consistent with ovulation and subsequent pregnancy forbehaviorally subordinate females living in groups with unrelated males or inwhich a reversal in female dominance status occurred. Results suggest thatin addition to changes in female reproductive endocrinology associated with
1Departments of Psychology and Biology, University of Nebraska at Omaha, Omaha, Nebraska.
2Department of Biology, University of Maryland, College Park, Maryland.
3Philadelphia Zoo, Philadelphia, Pennsylvania.
4Current address: Department of Psychiatry, University of Illinois at Chicago. 5To Whom Correspondence should be addressed; e-mail: jfrench@unomaha.edu.
0164-0291/03/1200-1281/0 C 2003 Plenum Publishing Corporation
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz puberty, the regulation of reproduction in females in wild callitrichid groupscan be sensitive to status and relatedness to breeding males.KEY WORDS: ovarian cycles, fecal steroids, seasonality, reproductive suppression. INTRODUCTION
In groups of cooperatively breeding mammals, reproduction is typi-
cally limited to a single, dominant female (Creel and Waser, 1997; French,1997; Moehlman and Hofer, 1997). In the callitrichid primates, a variety ofmechanisms can produce this dramatic reproductive skew among females. The mechanisms include suppression of ovulation and ovarian production ofsteroid hormones in older subordinates (Abbott and George, 1991; Carlsonet al., 1997; French et al., 1984; K ¨uderling et al., 1995; Saltzman et al., 1997a;Savage et al., 1988; Smith et al., 1997) and behaviorally-mediated mecha-nisms such as agonistic interactions or inhibition of sexual behavior (incestavoidance; French and Inglett, 1989, 1991; Saltzman et al., 1997b; Ziegler andSousa, 2002). Most studies of the proximate mechanisms underlying repro-ductive skew in callitrichid social groups have been conducted under captiveconditions. It seems likely that the rules regulating reproductive function insubordinates might operate differently under field conditions, where energyconstraints, e.g., foraging and food acquisition, travel, and predator avoid-ance, are more likely to shape reproductive decision-making. Under theseconditions, resources to support reproduction by dominant breeders mightbe limited, or reproductive attempts by subordinates may be more risky thanunder more benign captive conditions. In either event, we might expect tosee more dramatic socially-mediated reproductive inhibition in wild than incaptive callitrichid primates. Conversely, without the constraint of enforcedproximity to dominant females that cage-housing produces, subordinate fe-males may be less likely to encounter potential inhibitory stimuli emanatingfrom dominant females, at least during active phases of the day, and mayalso be more likely to encounter stimulatory cues from unrelated males thatare dispersing or in adjacent groups (Digby, 1999; Digby and Ferrari, 1994;Lazaro-Perea, 2001). Under these conditions, then, we might expect lesspronounced inhibition among subordinate females.
The development of noninvasive hormone monitoring methodologies
(Whitten et al., 1998) has allowed reproductive biologists to address ques-tions regarding endocrine mechanisms of reproduction in contexts in whichcomplex ecological and demographic factors have full play. Indeed, some im-portant differences in the regulation of reproduction in callitrichid primateshave been revealed in preliminary studies on cotton-top tamarins (Saguinusoedipus). In captive settings, daughters and subordinate females in social
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
groups of cotton-top tamarins are universally anovulatory while in an intactfamily group (French et al., 1984; Savage et al., 1988; Ziegler et al., 1987). However, a recent study on a single group of wild tamarins indicated that alldaughters in the group (n = 3) showed signs of ovarian activity as reflected inperiodic elevations in fecal progesterone and estrogen metabolites (Savageet al., 1997). These results confirm our suspicion that the regulation of re-production in females in wild callitrichid groups may be more complicatedthan we would expect based on results from captive animals.
In captive golden lion tamarins, details of ovarian function in adult fe-
males have been well-established via noninvasively collected urine samples. Nonconceptive ovarian cycle length is 19.6 d, and endocrine profiles suggesta gestation length of 125 d (French et al., 2002; French and Stribley 1985). Incontrast to most other callitrichid primates, subordinate females and daugh-ters in captivity exhibit no social suppression of ovarian function, as assessedby excreted urinary steroids, past the age of puberty (∼ 14–16 mo: Chaouiand Hasler-Gallusser 1999; De Vleeschouwer, et al., 2000; French et al., 1989;van Elsacker et al., 1994).
We evaluated the potential to assess reproductive function in wild lion
tamarins via fecal steroid monitoring. We tested whether the measures coulddocument changes in reproductive status, e.g., pregnancy, in dominant breed-ing females. Further, we examined the reproductive status of several subor-dinate females—daughters living in natal groups and subordinate femalesresiding in groups with unrelated dominant breeders—for a preliminary as-sessment of the impact of social environments on ovarian activity. We moni-tored reproductive endocrinology in dominant breeding females and daugh-ters/subordinates in multiple social groups during 4 breeding seasons (1996through 1999). The results demonstrate the utility of fecal steroid hormonemonitoring in the species and in the context of wild tamarins, and confirmthe complex seasonal, maturational, and social influences on reproductionin the golden lion tamarin. Focal Subjects and Study Site
We conducted all observations and sample collections at the Reserva
Biol ´ogica de Po¸co das Antas, a 6300 ha reserve in southeastern Rio de JaneiroState, Brazil. The subjects were 10 dominant breeding females and 9 femalesthat were natal daughters or behaviorally subordinate females residing ingroups that contained a dominant, breeding female (Tables I and II). Theyresided in 9 different social groups. Exact birthdates are known for mostfemales in the study because the groups are under intensive observation
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz Table I. Dominant reproductive females sampled in study
throughout the year (Dietz et al., 1994; Dietz and Baker, 1993). For indi-viduals that entered study groups from outside the marked population, weestimated age based on weight (for young individuals) and on tooth wear(for older animals). We determined intra-group dominance status amongfemales from the incidence and direction of archwalks and chases, whichindicate dominant status, and mounts, which indicate subordinate status, inthe long-term behavioral records on the groups.
We collected samples from breeding females during a variety of phases
of the annual reproductive cycle, including between and throughout preg-nancy. Although we collected samples throughout the year, we focused oncollecting feces from breeding and subordinate females during the breeding
Table II. Subordinate females sampled
∗Intact Natal Group = female resides in family with suspected mother and father present in
group; Nonintact Natal Group = female resides in natal group with replacement breeder, male;Nonnatal Group = female has dispersed from natal group and joined group with suspectednonrelatives
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
season—mating, pregnancy, parturition, and lactation—from late Maythrough early February. We sampled one subordinate female in 1996, butthe remainder of the samples are from 1997, 1998, and 1999. Sample Collection and Storage
Observers located radio-collared groups early in the morning, before
emergence from the sleeping site. When tamarins left the sleeping sites,observers closely watched them for signs of defecation. Since concentrationsof fecal progesterone metabolites (but not estrogen metabolites) vary insamples collected in the morning and afternoon in other callitrichid primates(Sousa and Ziegler, 1998), we attempted to restrict sample collection tomornings. We collected by 0900 h 50% of samples, and >80% by 1200 h. Weincluded only samples that were unambiguous with regard to the animal oforigin in the analyses. We collected a single sample from an individual onany given day. We placed the feces in a plastic whirl-pak sample bag, froze itwithin 3–6 h of collection, and stored it at −18◦C until shipment to the U.S. ondry ice. Tests with fecal samples collected from captive callitrichids indicatedthat a delay in freezing fecal samples of up to 96 h post-collection did notalter progesterone metabolite concentrations >10%, relative to samples thatwere frozen immediately (Brewer and French, unpublished data). Sample Extraction and Assay
After thawing at room temperature we placed 0.125 g fractions of each
sample in round-bottom 20-mL test tubes. We removed seeds, leafy mate-rial, and insect parts from the fecal sample before weighing. We added a5-mL volume of solubilizer (40% methanol: 60% phosphate buffered saline(PBS)) and we placed the tubes on a shaker rack overnight (12–16 h). Wepelleted particulate matter in the resulting slurry by centrifugation for 15 minat 2000 g, and separated the supernatant into a clean test tube. Recoveryof tritiated steroid after extraction was 77 ± 3.3%. We refroze and storedextracted samples before assay. Enzyme Immunoassays (EIA)
. We assessed fecal pregnanediol-3-glucuronide (PdG) levels via an EIA.
The protocol was modified from French et al. (1996) as follows. We further
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
diluted extracted fecal samples with assay buffer (1:5) before measurementin order to place the sample concentration within the range of the standardcurve. We constituted PdG standards (Sigma Chemical Co., St. Louis MO)in 1:5 extraction buffer:PBS, and they ranged from 10,000 to 78 pg/wellin halving concentrations. We determined the precision of the fecal EIAsvia a fecal quality control pool. The fecal quality control pool consisted ofthe supernatant of extracted fecal samples from several different females. Intra-assay coefficients of variation for high and low concentration qualitycontrols were 10.2% and 6.8%. Interassay CVs were 11% and 14.4% for thehigh and low concentration pools, respectively. Serial dilutions of extractedfeces from females at different times during pregnancy and PdG standardsproduced parallel displacement curves (t(8) = 0.85, p > 0.40). We determinedthe accuracy of the fecal PdG EIA by spiking the supernatant of an extractedfemale lion tamarin fecal sample with 2000 pg of PdG standard. The recoveryof PdG was 108.6 ± 5.9% (n = 6).
We also measured fecal estrone conjugates (E1C) by an EIA. The E1C
EIA was described by French et al. (1996), and we modified it as follows. We further diluted extracted fecal samples with assay buffer (1:20) beforemeasurement in order to place the sample concentration within the range ofthe standard curve. We diluted E1C standards (estrone-3-glucuronide, SigmaChemical Co., St. Louis MO) in a 1:20 dilution of extraction buffer, and theyranged from 400 to 6.2 pg/well in halving concentrations. We determinedthe precision of the E1C EIAs via the fecal quality control pool. Intra-assayCVs for high and low concentration pools were 4% and 3.6%. InterassayCVs based on the same pools were 22.1% and 19.6%, respectively. Serialdilutions of extracted feces from females at different times during pregnancyand E1C standards produced parallel displacement curves (t(8) = 0.91, p >0.30). We determined the accuracy of the assay by spiking the supernatantof an extracted female lion tamarin fecal sample with 25 pg of E1C standard. The recovery of E1C standard was 96.0 + 2.7%(n = 6). Normative Endocrine Function in Breeding Adult Females
Fecal steroid analyses on samples (n = 440 samples, 10 females) from
breeding females in the Po¸co das Antas population revealed clear signs ofovulation, conception, and the onset of pregnancy. Figure 1 portrays repro-ductive profiles in adult females, showing the onset of ovarian function inJune and July, with sustained and elevated hormone concentrations duringpregnancy and declines in hormone concentrations following parturition.
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins Fig. 1. Fecal steroid profiles for two dominant, breeding females in different social groups in Po¸co das Antas, across different reproductive phases. Vertical dotted lines represent dates of parturition.
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz Table III. Means (±standard errors) for fecal pregnanediol glucuronide (PdG) and estro- gen conjugates (E1C) for dominant reproductive females while non-pregnant and during each
trimester of pregnancy (n = number of females sampled during the period)
1052.32 ± 172.82 2086.98 ± 677.86
2905.18 ± 1082.22 4467.64 ± 884.93
1Pregnancy status was confirmed by back-dating from the day of parturition, assuming a 126 d
2Trimesters were defined as three successive 42 d periods during pregnancy, back-dated from
The duration of elevated steroid concentration for both females is ca. 4 mo. Table III lists mean values for PdG and E1C for non-pregnant females andfemales in each trimester of pregnancy. Although fecal sampling collectionoccurred less frequently outside the main breeding period, the low con-centrations from February through June/July suggest a seasonal period ofovarian quiescence. Concentrations of both PdG and E1C during pregnancytended to be above >5000 ng/g feces, and reached levels as high as 10,000 –15,000 ng/g (PdG) and 5,000 to 7,000 ng/g (E1C). Pregnant females (dom-inant and subordinate) had significantly higher concentrations of estrogenand progesterone metabolites than when they were not pregnant, by at leastone order of magnitude (F(1, 11) s > 23.70, p s < 0.001). Comparison of Endocrine Function in Dominant and Subordinate Females
Social status clearly influenced levels of ovarian hormone excretion in
nonpregnant females. Figure 2 presents mean concentrations of PdG andE1C measured in fecal samples collected from 9 subordinate females and 10dominant breeding females in varying stages of reproduction. We contrastedmean concentrations of hormones in all samples from nonpregnant females(Fig. 2a and b) and from pregnant dominant and subordinate females (Fig. 2cand d) as a function of their status in the group: reproductively inactive sub-ordinate, reproductively active subordinate, and dominant female. Levels ofPdG excretion are significantly lower in subordinate females than in domi-nant females (F(2, 15) = 4.76, p < 0.02), regardless of whether subordinatesultimately became pregnant or not. While levels of E1C tended to be lowerin subordinates than in dominant females, the differences are not significant(F(2, 15) = 1.55, n.s.). Pregnancy status of females was clearly reflected infecal concentrations of PdG and E1C, (Fig. 2c and d). Pregnant subordi-nates had PdG concentrations that do not differ significantly from those of
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins Fig. 2. Mean concentrations of fecal E1C and PdG in females as a function of reproductive and social status in the group (anovulatory and ovulatory subordinates vs. dominant adult females (a and b) and for pregnant females that varied in social status (c and d). Note the different scales for the Y-axes on panels 3c and d. Anov Sub = noncycling subordinate female; Ov Sub = ovulatory subordinate; Ad Fem = dominant breeding female; Preg Sub = pregnant subordinate female; Preg Dom = pregnant dominant female.
dominant females (F(1, 11) = 2.11, n.s.). However, levels of E1C are signif-icantly higher in pregnant subordinates than levels in pregnant dominants(F(1, 11) = 6.33, p < 0.03). Patterns of Fecal Steroid Excretion in Subordinate Females and Daughters
We collected 249 samples from 9 females identified as daughters or
socially subordinate to a dominant breeding female. The females providedmultiple samples (range = 5 − 77, median number of samples = 33.5), andwe could therefore evaluate the resulting hormonal profiles qualitatively andquantitatively for signs of ovulation or pregnancy. In 5 females, there wasclear evidence of anovulatory status during part or all of the sampling period. Figure 3 presents the profiles for 4 of them. Two of them (#685, #688, Fig. 3a,3b) were 1-yr-old daughters living in intact natal groups, and most samplescollected during this period had hormone concentrations at or just above theminimal level of sensitivity of the assays. Female #723 (Fig. 3c) had emigrated
International Journal of Primatology [ijop]
Fig. 3. Nonconceptive fecal steroid profiles for daughters and subordinate females in the presence of mothers or behaviorally-dominant females. For the duration of most of the sampling period for each female, there is little evidence for ovarian activity and/or pregnancy. See Table I and text for details on each female.
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
from her natal group as a 1-yr-old with her behaviorally-dominant sister(#689;) and joined a new group. Concentrations of fecal ovarian steroids for#723 never reached levels suggesting ovulatory function or pregnancy duringthe 1999 breeding season (Fig. 3c). Finally, there was no sign of ovarianactivity for #593 (Fig. 3d), a 2-yr-old daughter living in an intact natal group.
Two females well past the age of puberty were apparently anovulatory
while residing in groups as subordinate females, but ovulated and becamepregnant when they became a dominant female in the group. Female #689(Fig. 4a) was sampled as both a 1- and 2-yr-old while in her natal familygroup. In both years (1997 and 1998), she showed no evidence of ovulatorycycles or pregnancy. Female #689 dispersed with a younger sister at ca. 26 moof age and joined a new group as a dominant female. Samples collected earlyin the breeding season revealed no sign of pregnancy in #689, though shemight have produced a singleton on 10/4/1999. We also noted a period ofsteroid excretion levels suggesting anovulation for #720, a 4-yr-old subordi-nate female in a group she joined gradually from January through July 1998(Fig. 4b). During the duration of her residence in the group as a subordinate(through February 1999), #720 showed low and acyclic concentrations ofovarian steroid excretion. However, her reproductive status changed whenher social status changed. In February 1999, the dominant female left thegroup, and in May 1999, #720 displayed quantitative and qualitative steroidexcretion profiles suggesting successful pregnancy. She delivered a set of twininfants on 9/28/99, which is 120 – 130 d after her hormone concentrationsbecame elevated (Figure 4b).
Finally, 2 subordinate females displayed evidence of normal ovarian
function, including conception and pregnancy while they resided in socialgroups in the presence of a dominant female. Their profiles are in Figure 5aand b. Female #650 (Fig. 5a) was a 2-yr-old daughter residing in her intact na-tal group with her mother, the oldest breeding female in the study population(16 yrs). Female #650 had steroid excretion profiles suggesting pregnancy,both in terms of the period of elevation (ca 130 d) and in the concentrationsof steroids (multiple samples > 3, 000 ng/g feces). The dominant breedingfemale gave birth to infants on 9/25/97, after the date of apparent conceptionfor #650. The dominant female died on 11/5/97, 5 weeks after the birth ofher infants. The normal age of weaning in wild lion tamarins is ca. 12 weeksof age, and #650 was a primary carrier of the infants and also provided themwith animal prey. She remained with the infants after her father and brotherleft the group in apparent search of a new mate. No infant born to #650 wasseen by field staff, though the drop in steroid excretion in December suggeststhat her pregnancy was maintained for a normal gestation period.
Figure 5b portrays fecal steroid profiles for #603, a 3-yr-old in a group
with her mother (#436) and multiple adult males that had entered the group
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz Fig. 4. Fecal steroid profiles in two females who displayed patterns suggesting infertility while residing as subordinates in social groups, but then exhibited an onset of reproductive function after changing groups (#689) and after changing status in a group (#720). Dotted vertical lines indicate estimated dates of conception for each female.
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins Fig. 5. Conceptive fecal steroid profiles for daughter residing in an intact family group (#650) and a subordinate female residing in her natal group with a replacement male (#603). Although the profile for #650 suggests pregnancy, no infants were observed for her. Dotted lines estimate the dates of conception, and solid vertical arrows indicate dates of parturition for female 603.
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
after 603’s father died. Female 603 became pregnant twice (1998 and 1999),carried twins to term, and successfully reared both sets of infants. Concur-rently her mother (#436, see Fig. 2a) also birthed and reared 2 sets of infants. Mother and daughter produced infants 30 days apart in 1997, and 10 daysapart in 1998. In both cases, #436 delivered her litter first. DISCUSSION
Our data clearly indicate that, as in other species (ungulates: Kapke
et al., 1999, carnivores: Creel et al., 1992, 1997; neotropical primates: Strierand Ziegler, 1997; cercopithecine primates: Wasser et al., 1988; great apes:Whitten et al., 1998) useful information regarding female reproductive func-tion can be acquired via the analysis of excreted steroid hormones in fecesin lion tamarins. The profiles generated from breeding adult females clearlyshowed that we can differentiate pregnant from nonpregnant females onthe basis of both qualitative and quantitative assessments. Daily collectionsunder the controlled conditions of captivity can yield profiles that are usefulto pinpoint the periovulatory phase of the ovarian cycle (French et al., 2002). Under the conditions and demands of sample collection in the field, we wereunable to collect samples with sufficient frequency to identify subtle detailsof the ovarian cycle during development (in subordinates) or during noncon-ceptive cycles (in breeders). However, the close agreement between periodsof elevated steroids in dominant focal females (Fig. 1) and estimates of ges-tation length in captive lion tamarins (French and Stribley, 1985; Kleiman,1978) clearly indicates the biological validity of the sampling schedule tomonitor events between conception and parturition.
Although details of ovarian steroid hormone metabolism and excretion
are known for urine in lion tamarins (French and Stribley, 1985), little isknown about the metabolic fate of ovarian steroids excreted in their feces. Knowledge of steroid clearance in lion tamarins is critical, since there isconsiderable intergeneric variability in gut transit time and steroid hormoneclearance rates among other callitrichid genera (Saguinus and Callithrix:Ziegler et al., 1996). Further, recent work on squirrel monkeys suggests thatestrogens and progestagens may be excreted in feces at different rates, withprogestagens clearing more rapidly than estrogens (Moorman et al., 2002). Knowledge of these parameters may increase our ability to resolve moresubtle details of steroid hormone profiles in wild lion tamarins. Neverthelessour data are comparable to levels of fecal steroids from other wild callitrichidprimates (cotton-top tamarins; Savage et al. 1997).
Dominant females differed from subordinates in several aspects of
endocrine function. Among nonpregnant females, both anovulatory and
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
ovulatory subordinates had significantly lower concentrations of PdG thanthose of dominant adult females, and subordinates tended to have lower lev-els of E1C than those of dominants. That anovulatory females have lowerconcentrations of ovarian steroids than dominants is not surprising; however,ovulatory subordinates also were clearly distinguishable from dominants onthe basis of steroid hormone levels. In other callitrichid species in whichsubordinates ovulated, ovarian hormone profiles in the subordinates arealso distinguishable from those of dominants, with lower peak levels of pro-gesterone or progesterone metabolites on both and a shorter luteal phaseduration (Saltzman et al., 1994; Smith et al., 1997).
Social status was not associated with differences in PdG concentra-
tions, in pregnant females. However, pregnant subordinates had significantlyhigher levels of E1C than those of pregnant dominant females. Subordinatefemales are likely attempting independent reproduction for the first time,while dominants varied in the number of previous litters, so it is possiblethat the differences do not reflect the influence of social status but insteadportray parity or age effects. However, several dominant primiparous fe-males in our sample displayed no unusually high E1C value. Although weattempted to sample evenly throughout pregnancy, it is also possible thatsamples from pregnant subordinates were more likely to be collected at atime when hormone concentrations are high, e.g., second trimester, thanat other times, but the lack of a difference in PdG concentrations arguesagainst this interpretation. In any event, high prepartum E1C is associatedwith traits associated with reduced reproductive performance. In pregnantwild lion tamarins, mothers with high prepartum E1C have offspring withlower neonatal weights (Bales et al. 2002) and elevated estrogen concen-trations are associated with lower maternal care and poor infant outcomein other callitrichids (Fite and French, 2000; contra Pryce et al., 1988). Sub-ordinate female lion tamarins that attempt reproduction have significantlylower reproductive success (estimated by the number of offspring surviving6 mo) than dominants (Dietz and Baker, 1993), and the effects of elevatedE1C on reduced fetal growth and reduced maternal care may contribute tothis effect.
Although our small sample of subordinate females limits our conclu-
sions the endocrine profiles generated from the data suggest that ≥ 3 fea-tures influence reproductive function in daughters and subordinate femalesin groups of wild lion tamarins. Among them are age (younger females arelikely to show patterns of infertility, older females are likely to show elevatedsteroid concentrations), composition of the group (older females in groupswith both biological parents are less likely to show elevated steroid con-centrations than females in groups with step-parents or no related adults),and relative dominance status among females (older females that are clear
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
social subordinates are less likely to display elevated steroid concentrations). It is difficult to disentangle the effects of age, nutritional status, and socialstatus. However, as is increasingly apparent from both theoretical analy-ses of reproductive suppression in cooperatively breeding species (Creeland Waser, 1991; French 1997) and elegant experimental work on captiveanimals (Saltzman et al., 1997a, 1997b; Widowski et al., 1990), reproductivesuppression in female callitrichids is a physiological response that is sensitiveto a complex suite of variables.
As in captive studies on other callitrichid primates, age was a significant
predictor of whether a subordinate female tamarin showed ovarian activity. Only one of the 4 females that were sampled when they were <16 mo oldshowed signs of elevated hormone excretion (#704), and she had only a singleelevated sample with no sign before or after of continued activity. In addition,she was one of the females whose birthdate was estimated rather than known;therefore, she might have been older than estimated. Data on reproductiveontogeny in captive female lion tamarins residing in intact family groups alsosuggest that females <14 mo old have low and acyclic patterns of urinaryestrogen excretion (French, 1997; French et al., 1989; French and Stribley,1985). In the Po¸co das Antas population, no female in her first year has everconceived offspring. Together, these observations suggest that the patternof low and acyclic concentrations of steroid excretion in 1-yr-old females isnot socially-mediated, but instead reflects a prepubertal state in the female. However, low and acyclic steroid excretion profiles in older females (>20 mo:#723, #593, #689, #720) probably reflect social influences on ovarian function.
Only one female (#650) conceived while residing in a natal group in
which no unrelated male was present. Her 16-yr-old mother was the oldestfocal female at Po¸co das Antas. Although the latter was pregnant and gavebirth during the sampling period, she was in poor health and died 5 weekspostpartum. Further, before her death she did not engage in the social roletypical of a dominant female, such as leading group movement and partici-pating in agonistic responses during intergroup encounters. Indeed, #650was among the most aggressive group members during the territorial en-counters. Recently, examples of subordinate reproduction in several speciessuggest that subordinate females are sensitive to the reproductive or healthstatus of the dominant female, and the onset of independent reproductionis based on these assessments. For example, in common marmosets, subor-dinate female ovulation was common (8 of 10) in family groups where themother was ovulating but did not produce viable offspring, but subordinatefemale ovulation was much less common (4 of 16) when the mother wasregularly conceiving and carrying infants to term (Saltzman et al. 1997a). Perhaps subordinate female lion tamarins are sensitive to the dominant’sreproductive potential, and are more likely to both ovulate and to conceive
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
when the dominant is old or anovulatory. Further data on subordinate fe-males in groups with aging or sick dominant females will help to address thispossibility.
Our endocrine data, together with long-term field records from Po¸co
das Antas (Baker et al. 2002) suggest that subordinate females may choosewhether to reproduce. Breeding records indicate that among 2-yr-old fe-males, not a single female (of 17) that was co-resident in a group only withrelated males (fathers and/or brothers) produced offspring, though our en-docrine data presented suggest that one female may have been pregnant. Incontrast, 3 of 8 2-yr-old subordinates that lived in groups with ≥1 unrelatedmales produced offspring. If subordinates continue to reside in intact natalgroups, the likelihood of pregnancy increases: 55% of 3-yr-old females, andall 4-yr-old females, became pregnant while living in groups that containedonly related males. There are a variety of mechanisms that could producethis age-graded choice among subordinate females. First, it may be that stim-ulation from unrelated males is more critical for the onset of reproductivefunction in younger females than in older females (French et al., 1984; Heis-termann et al., 1989; Widowski et al., 1990), and thus older subordinates com-mence ovulation in the absence of unfamiliar males. Secondly, older femalesmay be more successful in seeking and achieving extragroup copulationsfrom neighboring males (Digby, 1999) than younger females are, and subse-quently have a higher rate of pregnancy. Finally, as females grow older andresidual reproductive value continues to decrease, subordinate females maybecome more tolerant of incestuous matings. This option may reflect a betterchoice for an older subordinate, which may run the risk of missing anotherbreeding opportunity. The benefits of this option are considerably enhancedwhen considered in light of the high costs of female dispersal (Baker and Di-etz, 1996). Knowledge about the paternity of offspring born to subordinatefemales would help differentiate among possible mechanisms. ACKNOWLEDGMENTS
We sincerely thank the many fieldworkers in Brasil who contributed
to the infrastructure of Po¸co das Antas and collection of samples, especiallyDenise Rambaldi, Reagan Lake, and the Ecolog´ıa Field Team. Kevin Brewerand Dr. Tessa Smith contributed to the development of fecal assays, and KimPatera and Jennifer McWilliams helped to process samples. The CallitrichidLab Research Group and Dr. Tessa Smith kindly provided useful commentson previous versions of the paper. The study and the field site was supportedby funds from the following sources: National Science Foundation (BCS97-27687 to JMD, IBN 97-23842, OSR 92-99225, and IBN 00-91030 to JAF,
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
predoctoral fellowship to KB; DBI 96-02266 RTG, University of Maryland);Sigma Xi, FONZ, and the University of Maryland. Permission to collect datain Brazil was provided by CNPq (Brazilian Research Council) and IBAMA(Brazilian Institute for the Environment), and importation of fecal samplesin the U.S. was conducted under the auspices of CITES, USDA, and CDC. REFERENCES
Abbott, D. H., and George, L. M. (1991). Reproductive consequences of changing social status
in female common marmosets. In Box, H. O. (ed.) Primate Responses to EnvironmentalChange, Chapman and Hall, London, pp. 295–310.
Baker, A. J., and Dietz, J. M. (1996). Immigration in wild groups of golden lion tamarins
(Leontopithecus rosalia). Am. J. Primatol. 38: 47–56.
Baker, A. J., Bales, K., and Dietz, J. (2002). Mating system and group dynamics in lion tamarins.
In Kleiman, D. G., and Rylands, A. B. (eds.), Lion Tamarins: Biology and Conservation,Smithsonian Press, Washington, DC, pp. 188–212.
Bales, K., French, J. A., and Dietz, J. (2002). Explaining variation in maternal care in a cooper-
atively breeding species. Anim. Behav. 63: 453–461.
Carlson, A. A., Ziegler, T. E., and Snowdon, C.T. (1997). Ovarian function of pygmy marmoset
daughters (Cebuella pygmaea) in intact and motherless families. Am. J. Primatol. 43:347–355.
Chaoui, N. J., and Hasler-Gallusser, S. (1999). Incomplete sexual suppression in Leontopithecuschrysomelas: A behavioural and hormonal study in a semi-natural environment. FoliaPrimatol. 70:47–54.
Creel, S., Creel, N. M., Mills, M. G. L., and Monfort, S. L. (1997). Rank and reproduction in
cooperatively breeding African wild dogs: Behavior and endocrine correlates. Behav. Ecol. 8: 298–306.
Creel, S. R., Creel, N. M., Wildt, D. E., Monfort, S. L. (1992). Behavioural and endocrine
mechanisms of reproductive suppression in Serengeti dwarf mongooses. Anim. Behav. 43:231–245.
Creel, S. R., Waser, P. M. (1991). Failures of reproductive suppression in dwarf mongooses
(Helogale parvula): Accident or adaptation? Behav. Ecol. 2: 7–15.
Creel, S. R., Waser, P. M. (1997). Variation in reproductive suppression among dwarf mongooses:
Interplay between mechanisms and evolution. In Solomon, N. G., and French, J. A. (eds.),Cooperative Breeding in Mammals, Cambridge University Press, Cambridge, UK, pp. 150–170.
De Vleeschouwer, K., Van Elsacker, L., Heistermann, M., and Leus, K. (2000). An evaluation of
the suitability of contraceptive methods in golden-headed lion tamarins (Leontopithecuschrysomelas), with emphasis on melengestrol acetate (MGA) implants: 2 Endocrinologicaland behavior effects. Anim. Welfare 9: 385–401.
Dietz, J. M., and Baker, A. J. (1993). Polygyny and female reproductive success in golden lion
tamarins, Leontopithecus rosalia. Anim. Behav. 46: 1067–1078.
Dietz, J. M., Baker, A. J., and Miglioretti, D. (1994). Seasonal variation in reproduction, juvenile
growth, and adult body mass in golden lion tamarins (Leontopithecus rosalia). Am. J. Primatol. 34: 115–132.
Digby, L. J. (1999). Sexual behavior and extragroup copulations in a wild population of common
marmosets (Callithrix jacchus). Folia Primatol. 70: 136–145.
Digby, L. J., and Ferrari, S. F. (1994). Multiple breeding females in free-ranging groups of
Callithrix jacchus. Int. J. Primatol. 15: 389–397.
Fite, J. E., and French, J. A. (2000). Pre- and postpartum sex steroids in female marmosets
(Callithrix kuhlii): Is there a link with infant survivorship and maternal behavior? Horm. Behav. 38:1–12.
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
French, J. A. (1997). Proximate regulation of singular breeding in callitrichid primates. In
Solomon, N. G., and French, J. A.(eds.), Cooperative Breeding in Mammals, CambridgeUniversity Press, Cambridge, UK, pp. 34–75.
French, J. A., Abbott, D. H., and Snowdon, C. T. (1984). The effect of social environment
on estrogen excretion, scent marking, and sociosexual behavior in tamarins (Saguinusoedipus). Am. J. Primatol. 12: 469–478.
French, J. A., Brewer, K. J., Schaffner, C. M., Schalley, J., Hightower-Merritt, D., Smith, T. E.,
and Bell, S. M. (1996). Urinary steroid and gonadotropin excretion across the reproductivecycle in female black tufted-ear marmosets (Callithrix kuhli). Am. J. Primatol. 40: 231–246.
French, J. A., De Vleeschouwer, K., Bales, K., Heistermann, M. (2002). Reproduc-
tive biology of lion tamarins. In Kleiman, D. G., and Rylands, A. B., (eds.), LionTamarins: Biology and Conservation, Smithsonian Press, Washington, DC, pp.133–156.
French, J. A., and Inglett, B. J. (1989). Female-female aggression and male indifference in
response to unfamiliar intruders in lion tamarins. Anim. Behav. 37: 487–497.
French, J. A., and Inglett, B. J. (1991). Responses to novel social stimuli in tamarins: A com-
parative perspective. In Box, H.O. (ed.), Primate Responses to Environmental Change,Chapman and Hall, London, pp. 275–294.
French, J. A., Inglett, B. J., and Dethlefs, T. M. (1989). The reproductive status of nonbreeding
groups members in captive golden lion tamarin social groups. Am. J. Primatol. 18: 73–86.
French, J. A., and Stribley, J. A. (1985). Patterns of urinary oestrogen excretion in female golden
lion tamarins (Leontopithecus rosalia). J. Reprod. Fertil. 75: 537–46.
Heistermann, M., Kleiss, E., Pr ¨ove, E., W ¨olters, H.-J. (1989). Fertility status, dominance, and
scent maring behavior of family-housed female cotton-top tamarins (Saguinus oedipus) inabsence of their mothers. Am. J. Primatol. 18: 177–189.
Kapke, C. A., Arcese, P., Ziegler, T. E., and Scheffler, G. R. (1999). Estradiol and proges-
terone metabolite concentrations in white-tailed deer (Odocoileus virginianus) feces. J. Zoo. Wildlife Med. 30: 361–371.
Kleiman, D. G. (1978). Characteristics of the reproductive and sociosexual interactions in pairs
of lion tamarins (Leontopithecus rosalia) during the reproductive cycle. In Kleiman, D.G,(ed.), The Biology and Conservation of the Callitrichidae, Smithsonian Press, Washington,DC, pp. 181–190.
K ¨uderling, I., Evans, C., Abbott, D. H., Pryce, C. R., and Epple, G. (1995). Differential excre-
tion of urinary oestrogen by breeding females and daughters in the red-bellied tamarin(Saguinus labiatus). Folia Primatol. 64: 140–145.
Lazaro-Perea, C. (2001). Intergroup interactions in wild common marmosets, Callithrix jacchus:
Territorial defence and assessment of neighbours. Anim. Behav. 62: 11–21.
Moehlman, P., and Hofer, M. (1997). Cooperative breeding, reproductive suppression, and
body size in canids. In Solomon, N. G., and French, J. A. (eds.), Cooperative Breeding inMammals, Cambridge University Press, Cambridge, UK, pp. 76–128.
Moorman, E. A., Mendoza, S. P., Shideler, S. E., and Lasley, B. L. (2002). Excretion and measure-
ment of estradiol and progesterone metabolites in the feces and urine of female squirrelmonkeys (Saimiri sciureus). Am. J. Primatol. 57: 79–90.
Pryce, C. R., Abbott, D. H., Hodges, J. K., and Martin, R. D. (1988). Maternal behavior is related
to prepartum urinary estradiol levels in red-bellied tamarin monkeys. Physiol. Behav. 44:717–726.
Saltzman, W., Schultz-Darken, N. J., Scheffler, G., Wegner, F. H., and Abbott, D. H. (1994). Social
and reproductive influences on plasma cortisol in female marmoset monkeys. Physiol. Behav. 56: 801–810.
Saltzman, W., Severin, J. M., Schultz-Darken, N. J., Abbott, D. H. (1997a). Behavioral and social
correlates of escape from suppression of ovulation in female common marmosets housedwith the natal family. Am. J. Primatol. 41: 1–21.
Saltzman, W., Schultz-Darken, N. J., and Abbott, D. H. (1997b). Familial influences on ovulatory
function in common marmosets (Callithrix jacchus). Am. J. Primatol. 41: 159–178.
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
Savage, A., Ziegler, T. E., and Snowdon, C. T. (1988). Sociosexual develoment, pair bond
formation, and mechanisms of fertility suppression in female cotton-top tamarins (Saguinusoedipus oedipus). Am. J. Primatol. 14: 345–359.
Savage, A., Shideler, S. E., Soto, L. H., Causado, J., Giraldo, L. H., Lasley, B. L., and Snowdon,
C. T. (1997). Reproductive events of wild cotton-top tamarins (Saguinus oedipus) in Colom-bia. Am. J. Primatol. 43: 329–337
Sousa, M. B. C., and Ziegler, T. E. (1998). Diurnal variation on the excretion patterns of fecal
steroid in common marmoset (Callithrix jacchus) females. Am. J. Primatol. 46: 105–118.
Smith, T. E., Schaffner, C. M., and French, J. A. (1997). Social and developmental influences
on reproductive function in female Wied’s black tufted-ear marmosets (Callithrix kuhli). Horm. Behav. 31: 159–168.
Strier, K. B., and Ziegler, T. E. (1997). Behavioral and endocrine characteristics of the reproduc-
tive cycle in wild muriqui monkeys, Brachyteles arachnoids.Am. J. Primatol. 42: 299–310.
van Elsacker, L., Heistermann, M., Hodges, J. K., de Laet, A., and Verhayen, R. F. (1994).
Preliminary results on the evaluation of contraceptive implants in golden-headed liontamarins, Leontopithecus chrysomelas. Neotrop. Primates 2: 30–32.
Wasser, S. K., Risler, L., and Steiner, R. A. (1988). Excreted steroids in primate feces over the
menstrual cycle and pregnancy. Biol. Reprod. 39: 862–872.
Whitten, P. L., Brockman, D. K., and Stavisky, R. C. (1998). Recent advances in noninvasive
techniques to monitor hormone-behavior interactions . Am. J. Phys. Anthropol, Supp. 27:1–23.
Widowski, T. M., Ziegler, T. E., Elowson, A. M., and Snowdon, C. T. (1990). The role of males
in the stimulation of reproduction function in female cotton-top tamarins, Saguinus o. oedipus. Anim. Behav. 40: 731–741.
Ziegler, T. E., Bridson, W. E., Snowdon, C. T., and Eman, S. (1987). The endocrinology of
puberty and reproductive functioning in female cotton-top tamarins (Saguinus oedipus)under varying social conditions. Biol. Reprod. 37: 618–627.
Ziegler, T. E., Scheffler, G., Wittwer, D. J., Schultz-Darken, N. J., Snowdon, C. T., and Abbott,
D. H. (1996). Metabolism of reproductive steroids during the ovarian cycle in two speciesof callitrichids, Saguinus oedipus and Callithrix jacchus, and estimation of the ovulatoryperiod from fecal steroids. Biol Reprod 54: 91–99.
Ziegler, T. E., and Sousa, M. B. (2002) Parent-daughter relationships and social controls on
fertility in female common marmosets, Callithrix jacchus. Horm. Behav. 42: 356–367.
COGNITIVE NEUROSCIENCE AND NEUROPSYCHOLOGYTime-dependent e¡ect of transcranial directcurrent stimulation on the enhancement ofSuk Hoon Ohna, Chang-Il Parkd, Woo-Kyoung Yooe, Myoung-Hwan Kof, Kyung Pil Choia,Gyeong-Moon Kimb, Yong Taek Leec and Yun-Hee KimaDepartments of aPhysical Medicine and Rehabilitation, Division for Neurorehabilitation, bNeurology, Stroke and Cerebrovascular Center, Sam
Treatment of Nursing Home– Acquired Pneumonia KYLE MILLS, PharmD, BCPS, and A. CHRISTIE NELSON, PharmD University of Wyoming School of Pharmacy, Laramie, Wyoming BRADFORD T. WINSLOW, MD, FAAFP, Swedish Family Medicine Residency Program, Littleton, Colorado KATHRYN LEE SPRINGER, MD, Greater Denver Infectious Diseases, Denver, Colorado Pneumonia is an important cause of morbidity and
 International Journal of Primatology [ijop]
International Journal of Primatology, Vol. 24, No. 6, December 2003 ( C 2003)
Endocrine Monitoring of Wild Dominant and
International Journal of Primatology [ijop]
International Journal of Primatology, Vol. 24, No. 6, December 2003 ( C 2003)
Endocrine Monitoring of Wild Dominant and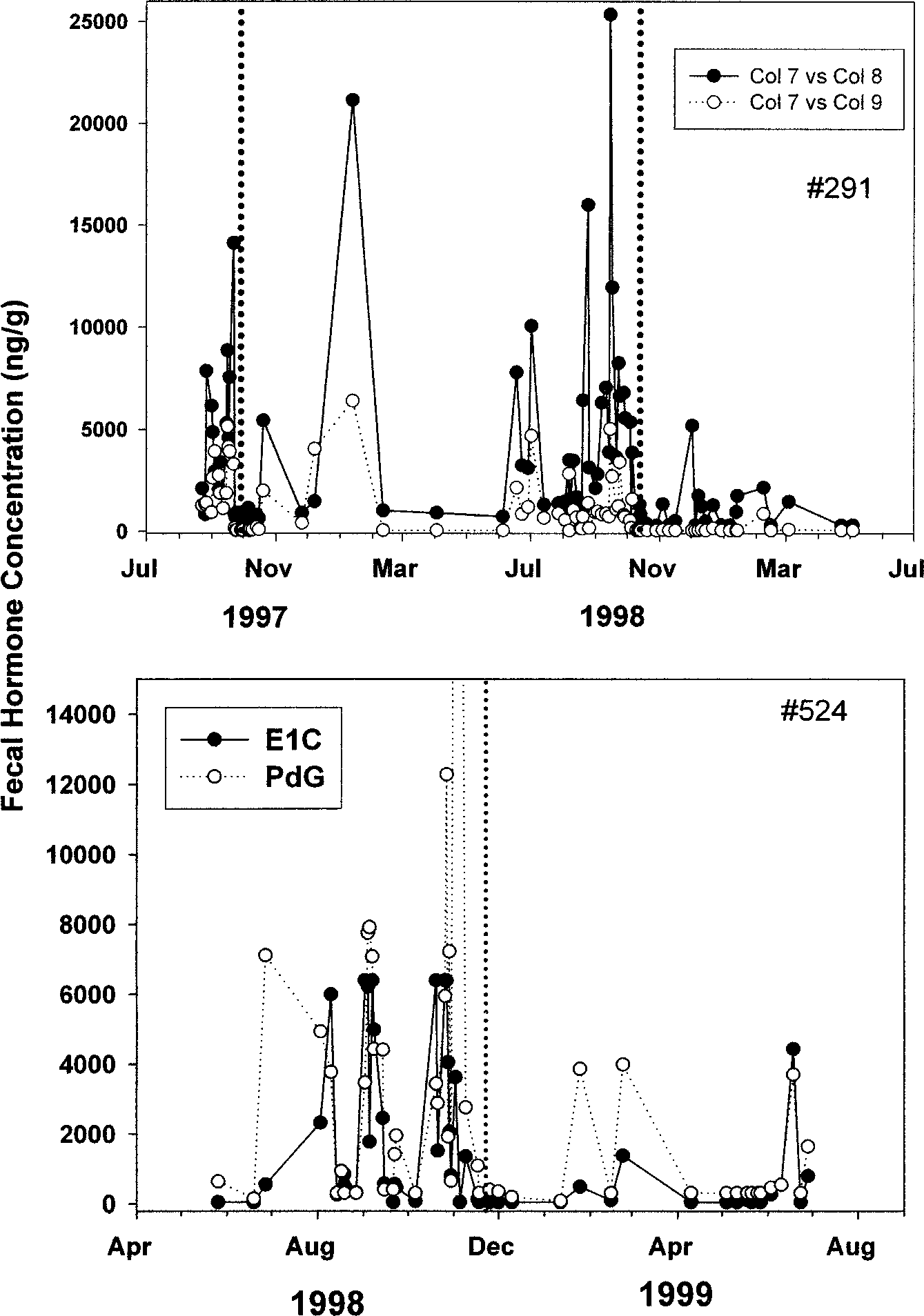 International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins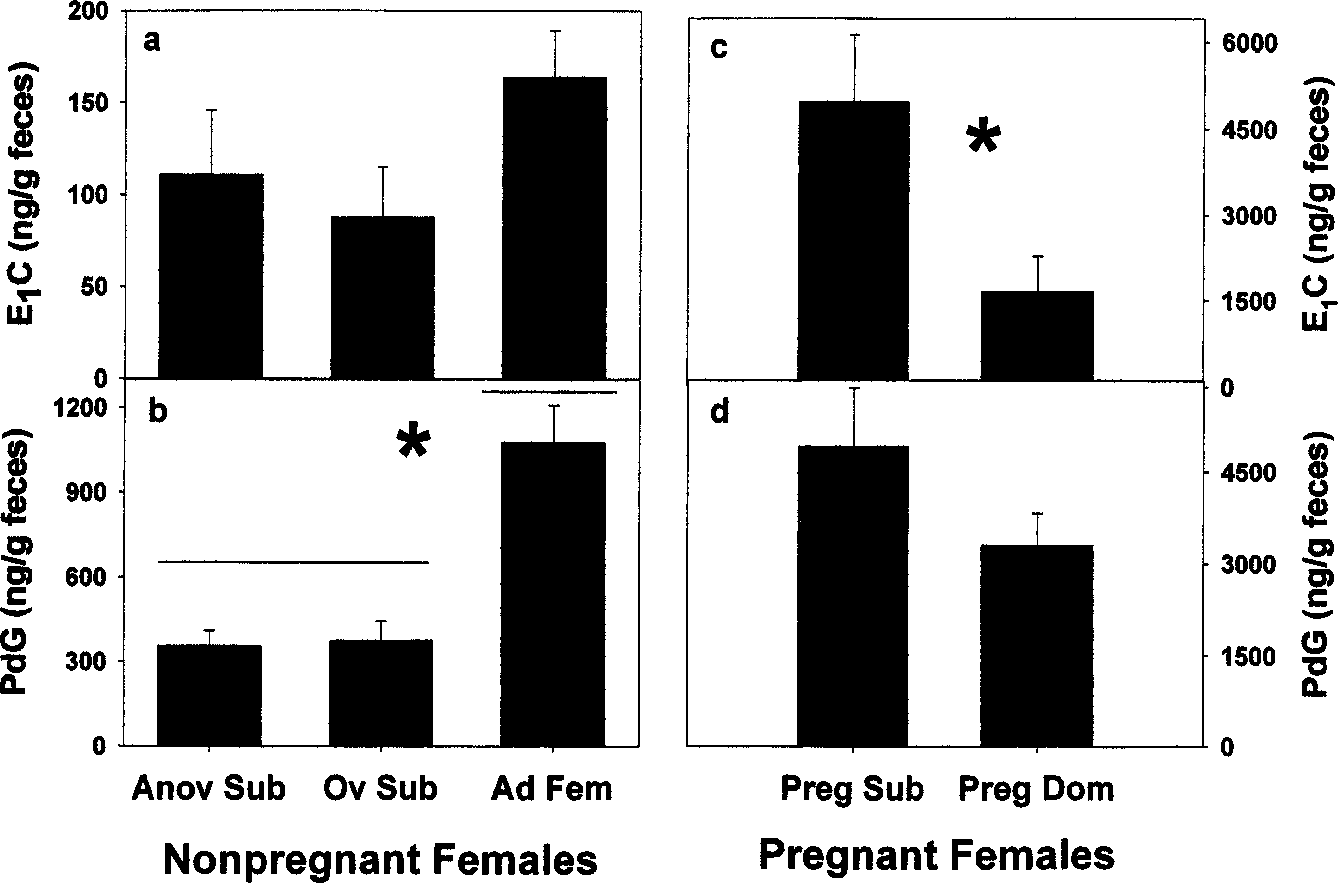 International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins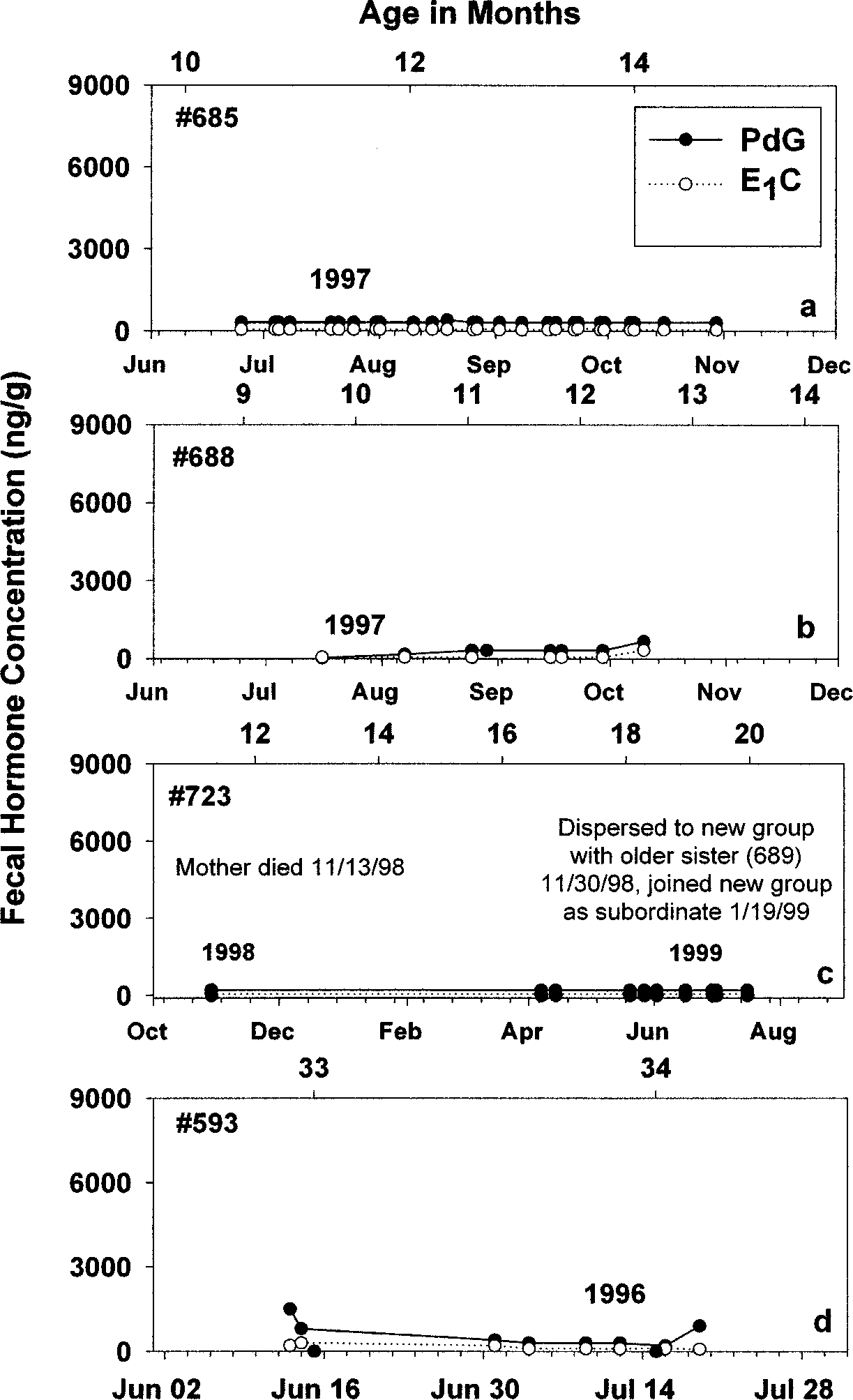 International Journal of Primatology [ijop]
Fig. 3. Nonconceptive fecal steroid profiles for daughters and subordinate females in
International Journal of Primatology [ijop]
Fig. 3. Nonconceptive fecal steroid profiles for daughters and subordinate females in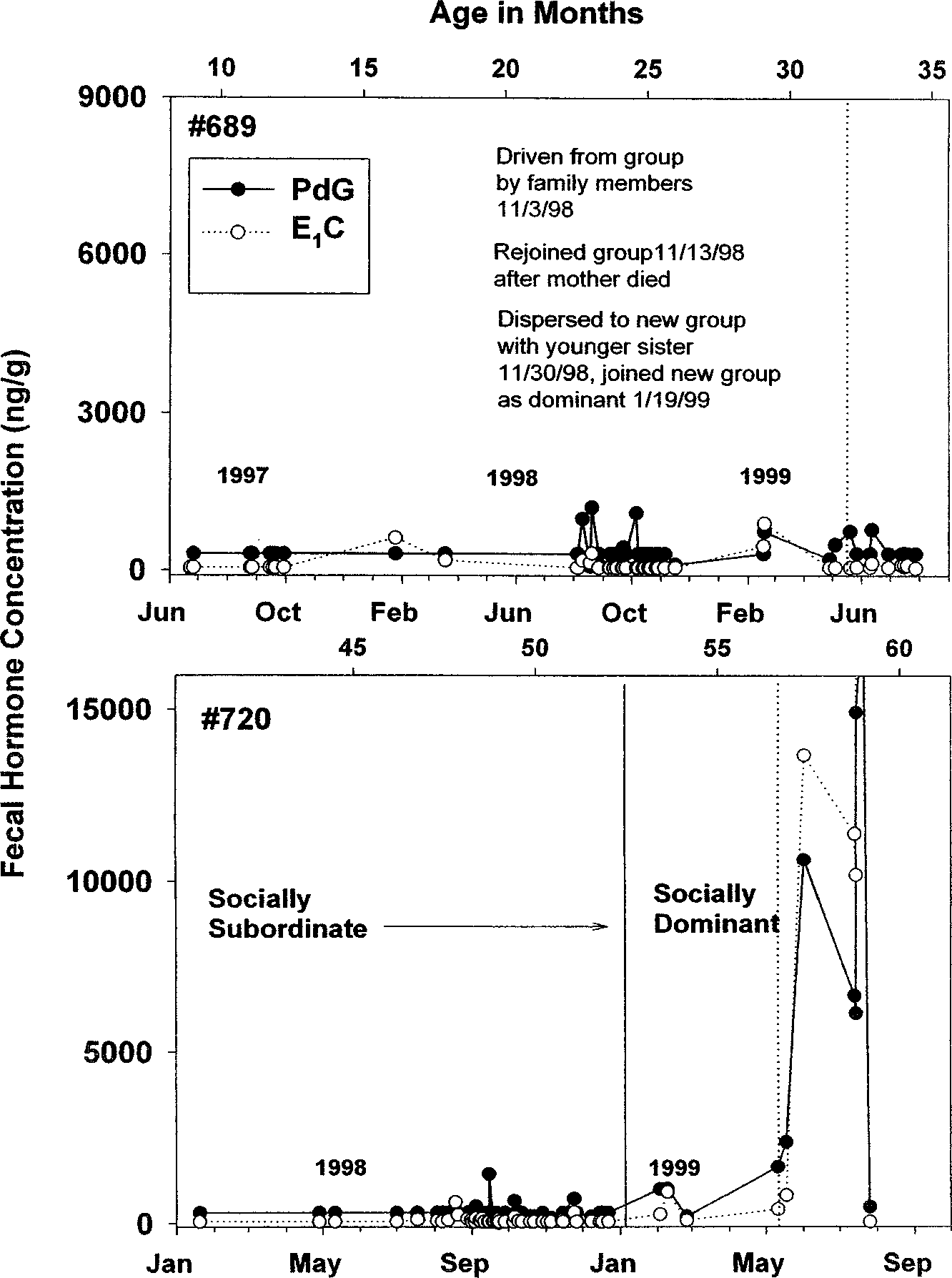 International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz
International Journal of Primatology [ijop]
French, Bales, Baker, and Dietz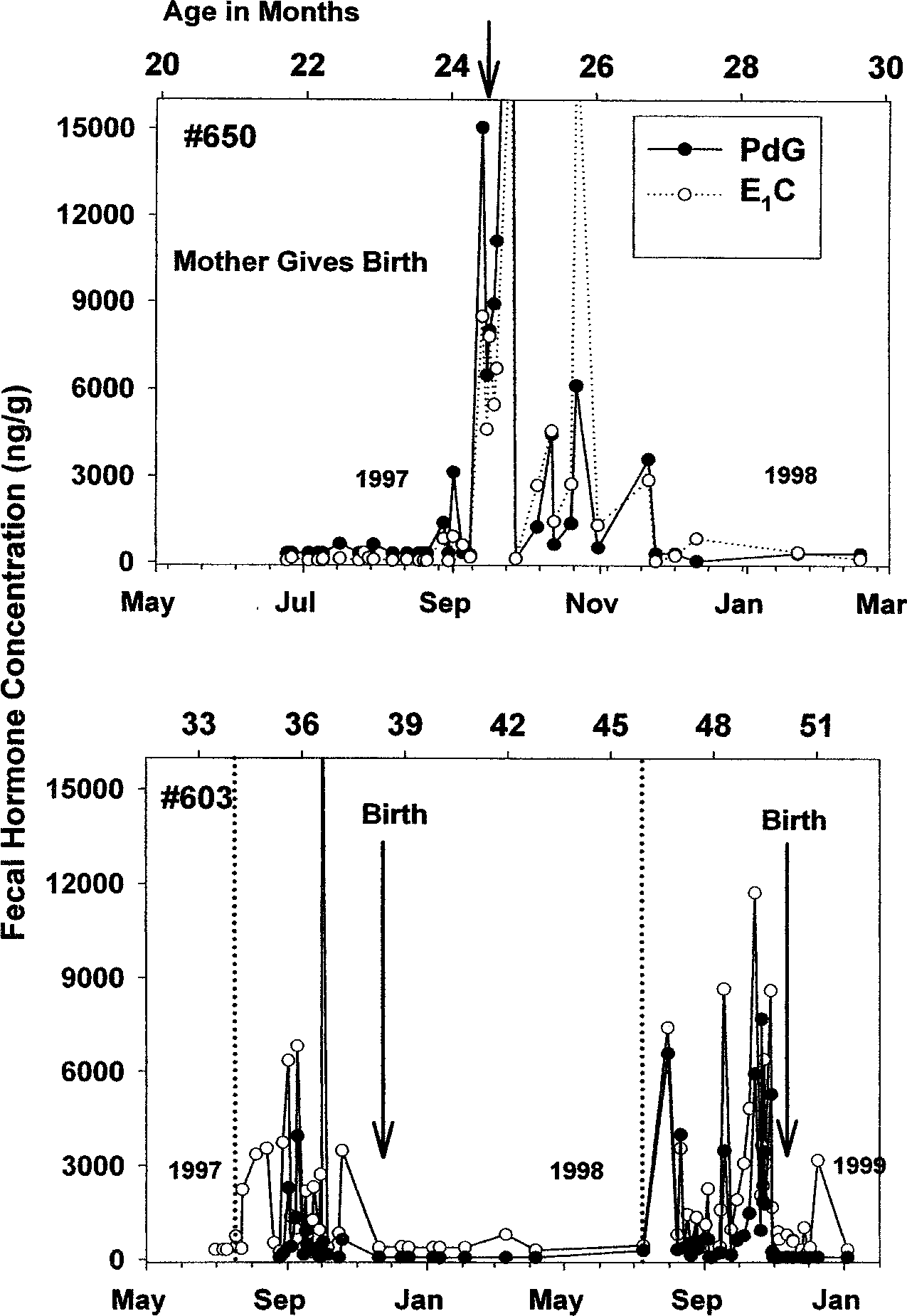 International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins
International Journal of Primatology [ijop]
Endocrinology of Wild Lion Tamarins