La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Evol_55_914.1902_1905.tp
Evolution, 55(9), 2001, pp. 1902–1905
CAN EXPERIMENTS ON CALORIC RESTRICTION BE RECONCILED WITH THE DISPOSABLE
SOMA THEORY FOR THE EVOLUTION OF SENESCENCE?
Deptartment of Biology, Leidy Laboratory, University of Pennsylvania, Philadelphia, Pennsylvania 19104-6018
A publication by Shanley and Kirkwood (2000) attempts to explain data on caloric restriction (CR) and
life extension in the context of the Disposable Soma (DS) theory for the evolution of senescence. As the authorsconcede, this juxtaposition appears at first to offend intuition: According to the DS theory, senescence is the resultof a tight budget for caloric energy, such that repair and maintenance functions are shortchanged; yet, in CR exper-iments, it is found that longevity decreases smoothly as the total caloric budget is increased. In the Shanley-Kirkwoodmodel, an optimized allocation of resources causes energy to be diverted away from somatic maintenance at a greaterrate than caloric intake increases, with the net result that more total energy is associated with less energy availablefor maintenance. In the present critique, the limitations of this model are detailed and its special assumptions reviewed. While the CR experiments find comparable life extension for males and females, measured relative to nonbreedingcontrols, the Shanley-Kirkwood model draws its energy budget from data on breeding females. In addition, the successin reproducing the observed relationship between feeding and longevity depends crucially on a mathematical rela-tionship between food availability and the probability of reproductive success which may be difficult to justify.
Aging, antagonistic pleiotropy, caloric restriction, dietary restriction, disposable soma, senescence.
prospects for successful reproduction and for survival ofyoung, vulnerable offspring is diminished; hence, it may be
According to the Disposable Soma (DS) theory (Kirkwood
advantageous to delay reproduction, devoting extra metabolic
1977), every living organism must budget its energy among
resources to maintenance and survival functions (Holliday
various priorities, including metabolism, growth, activity,
1989; Austad 1995; Masoro and Austad 1996). This theo-
and reproduction. Repair of damaged proteins and error-
retical framework creates a context for modeling the effect
checking in DNA replication constitute demands on energyresources that must compete with these other functions, and
with optimization of reproductive value; allocation of an un-
so their allocation is subject to compromise. Failure to do a
specified, scarce metabolic resource may be directed either
perfect job in these and other repair and maintenance func-
toward the immediate goal of reproduction or toward the
tions leads to an accumulation of damage over the lifetime
long-term benefit of forestalled senescence. In times when
of the organism, the outward manifestation of which is the
the food supply is stressed, the optimum balance may sensibly
constellation of symptoms we identify as senescence.
The DS theory for the evolution of aging is elegant and
In the DS theory, the scarce resource is specified to be
economical, with strong common-sense appeal. Perhaps for
caloric energy itself; this puts an added burden on the model,
this reason, it has enjoyed success relative to other theories
which must account for an increasing repair/maintenance
of senescence, in a field where experimental evidence is sub-
budget even as the overall caloric intake is declining steadily.
Perhaps for this reason, it was not until just last year that anattempt to reconcile DS with the CR data was first put for-
Caloric Restriction and Life Extension
ward. This is the model of Shanley and Kirkwood (2000).
One experimental area which is comparatively unequivocal
is the data on caloric restriction (CR). Laboratory animalsmaintained on a diet restricted in calories live longer than
The Shanley-Kirkwood (S-K) model is based in a detailed
ad libitum fed animals, and many of the physiological mark-
accounting of optimized energy rationing in times of famine
ers of senescence appear on a delayed schedule. These data
and times of plenty. Life history optimization is used to de-
pose a particular dilemma for the DS theory, with its foun-
termine the allocation of caloric resources, presumably under
dation in the metabolic energy budget. If senescence is caused
genetic control, between reproduction and maintenance of
by a failure to allocate adequate caloric energy for mainte-
the soma. Reproduction is energetically expensive for the
nance of the soma, why is it that the availability of caloric
females modeled exclusively in the paper, but it offers an
energy in abundance actually leads to more rapid senescence
immediate payoff; energy expended in maintenance affords
than occurs when energy is in short supply?
no payoff until much later, delaying the effects of accumu-
Theorists have explained the evolutionary provenance of
lated damage which manifest as reduced fertility and in-
the CR effect in terms of a shifted balance between the rel-
creased mortality. Under the model’s base assumptions, the
ative value of immediate reproduction and long-term surviv-
authors report that increasing food supply makes more energy
al: in times of famine, signaled by a meager food supply, the
available for reproduction, while the maintenance portion
holds constant over a wide range of caloric intake; hence,
Limitation of the Model to Breeding Females
the essence of the CR response remains unexplained. To en-
Half of one sentence justifies limitation of the S-K model
gineer an agreement with reality, the model is varied with
to females: ‘‘By common convention, the analysis is restrict-
ed to females, female demographic dominance being assumed
The first is a reproductive overhead, the cost of maintaining
. . . ’’ (Charlesworth 1994, p. 4). The reference is to the fact
fertility independent of whether a litter actually ensues. When
that population genetic analysis may be confined to females
reproductive overhead is introduced, Shanley and Kirkwood
with little loss of generality, and predictions concerning
find, after optimizing energy allocation, that reproduction
growth and steady-state population levels for mixed popu-
turns off abruptly at a threshold CR level. The implication
lation levels are little affected by male life histories. But this
for senescence is that there is a narrow range around this
is no reason to bypass males in the application of theories
threshold in which energy allocation for maintenance de-
of senescence or of CR. Life histories of males are separately
creases sharply with increasing food supply. This result has
optimized for fitness in a process analogous to that for fe-
the right sign to explain the data, but the curve is the wrong
males (Charlesworth 1994, p. 231), though, of course, the
shape. In experimental findings, longevity decreases smooth-
parameters of the cost/benefit calculus are different. There is
ly with increasing food availability over a wide range (Wein-
no logical basis in the Shanley-Kirkwood model for restrict-
The model is extended with a second parameter, which
The level of caloric restriction in lab settings as conven-
relates the probability of infant survival to ambient food sup-
tionally reported is measured relative to the ad libitum intake
ply. It is proposed that foraging results for the mouse in the
of nonbreeding animals, male and female. However, the CR
wild are a linear measure of the local availability of food;
variable in the S-K analysis is calibrated with respect to in-
thus restricted caloric intake is a proxy for ambient famine
take for breeding females. Pregnant and lactating females
conditions. The infant’s prospects for survival through the
consume up to twice as much food under ad libitum condi-
nursing period are assumed to increase linearly with ambient
tions (Bronson 1989). Without this large reservoir to draw
food availability; combined with the assumed linear rela-
down in times of caloric restriction, no adjustment of param-
tionship between caloric intake and fertility, this amounts to
eters in the model can reconcile it with the basic finding of
a quadratic dependence of the effective reproductive rate on
life extension due to caloric restriction, and, in fact, the op-
caloric intake. Under these assumptions, Shanley and Kirk-
posite result is predicted. It is an unstated prediction of the
wood report that optimization can reproduce a smooth re-
S-K model that breeding females should have a lifespan much
lationship between longevity and food intake which agrees
shortened compared to either males or nonbreeding females,
qualitatively with the experimental findings.
and this is not observed. It is only from this shortened base-line that their model is able to predict life extension. Shortcomings of the Shanley-Kirkwood Model
Fertility is reduced but not completely curtailed in male
Weaknesses of the model fall in three classes: First, there
mice, even under severe calorie restriction (De Paolo 1993).
is a mismatch in breeding status between animals in the model
The male control, fed ad libitum, consumed only half as much
and in the laboratory. The model assumes 50% of total met-
energy as a pregnant female to begin with. This male mouse
abolic energy allocated to reproduction in the ad libitum con-
on 50% caloric restriction still had most of its fertility (Merry
trols, and this figure is only credible for pregnant or lactating
and Holehan 1981), and weighed 60% as much as the control;
females. But the laboratory experiments in which the CR
yet it was more active (McCarter 1993), supported a stronger
effect is observed typically compare nonbreeding animals,
immune system (Venkatraman and Fernandes 1993), and
male and female, with their CR counterparts. The model ap-
lived one-third longer than the control (Weindruch and Sohal
plied to males and to nonbreeding females unequivocally
1997). Where did the energy come from to fuel its increased
predicts declining life span with declining caloric intake—
Second, there are questionable assumptions in the for-
Questionable Model of the Relationship Between Food
mulation of the model: Among several variations in the mod-
el, the only one that generates the observed qualitative be-
As described above, Shanley and Kirkwood report several
havior in the curve of life extension versus feeding level relies
versions of the model results, only one of which reproduces
upon a quadratically increasing relationship between the am-
an allocation of food energy to maintenance which gradually
bient food availability and the number of offspring success-
decreases with increasing food supply, supporting results in
fully weaned. This is justified with a kind of double counting,
agreement with observation. The key element in the model
that determines this behavior is the assumed relationship be-
Third, there are disparities between the model results and
tween ambient food supply and the probability of weaning
the CR data: The model can explain the inverse relationship
between food intake and lifespan only over a limited range
In this version of the model, food intake is taken as a proxy
of feeding levels; in experimental results, the relationship
variable, predicting the availability of food in the environ-
continues a smooth, quasi-linear behavior over more than a
ment ten weeks in the future, when offspring conceived as
factor of two, from the threshold of starvation at the low end
a result of present fertility impose their greatest caloric de-
to obesity at the high end (Weindruch et. al. 1986; Ross and
mand on the nursing mother. The infants’ prospects for sur-
vival through the nursing period is modeled as increasing
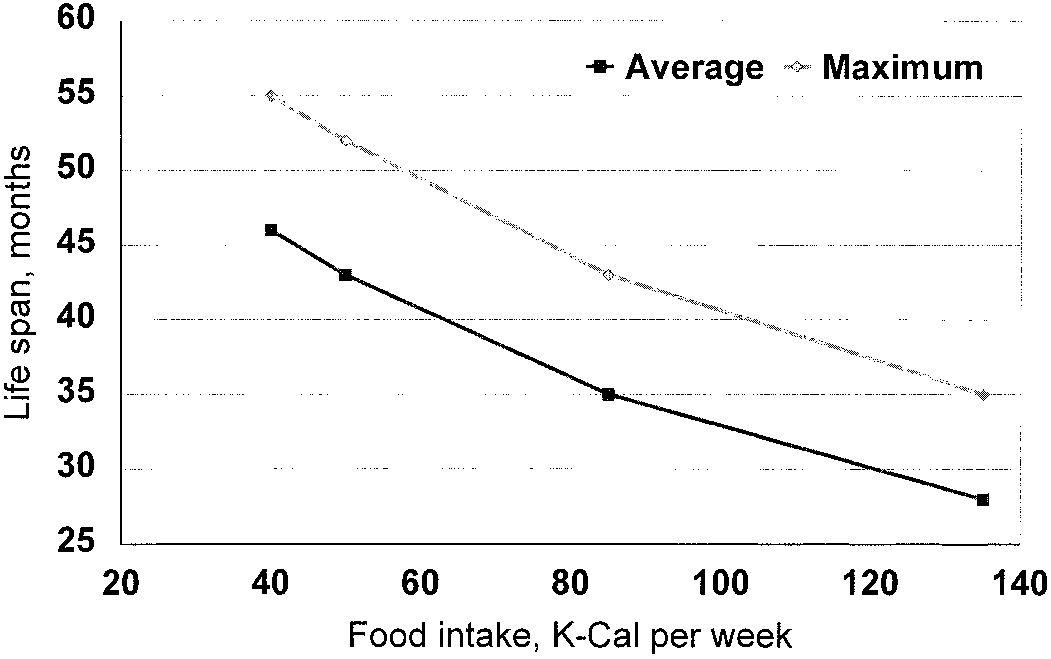
Life span of nonbreeding female mice increases smoothly
over a broad range as calories are reduced. Note that the increase
Results of the best-fitting version of the Shanley and Kirk-
continues substantially below the restriction level (80 K-cal) at
wood model. Energy that is not allocated to reproduction is con-
which female fertility is zero. This is difficult to reconcile with any
sidered available for somatic maintenance. Graph is for reproducing
model in which life-span extension derives from an energy reservoir
females. Note that the plot should not be directly compared to Figure
which is freed up as the animal diverts resources that had been
1 for this reason, and because both axes here have been normalized
devoted to reproduction. Graph adapted from Weindruch and Sohal
to body mass, which also varies with food intake. Graph adapted
1997, based on data from Weindruch et al. 1986.
linearly with ambient food availability, even as the mother’s
food supply support a successively larger increment in the
milk output per offspring is held constant.
In the base model, before this extra parameter is incor-
porated, there is already a linear per-offspring energy cost of
Ends of the Model’s Parameter Range
reproduction, based upon published laboratory measurements
Even the most successful variant of the S-K model cor-
(Millar 1987; Bronson 1989). But in the variant model, each
responds only to a limited range of the data for life extension
of these offspring is assigned a probability of survival which
from CR. The ends of the parameter ranges explored in their
is scaled by a second factor of caloric intake/ad libitum. Thus,
paper pose a difficulty, because the model has been optimized
in this variant, the relationship between surviving offspring
to work within these limits, and it falls apart at the bound-
and available calories becomes quadratic. Without this qua-
aries. There is a threshold feeding level at which fertility falls
dratic dependence, the model fails to reproduce the quali-
to zero. Below that level, somatic maintenance is predicted
tative characteristics of life extension via CR.
to drop sharply with further caloric restriction. But in CRexperiments, lifespan continues to increase smoothly even at
How the Shanley-Kirkwood Model Arrives at a
the lowest feeding levels (Weindruch et al. 1986) (see Figs.
Many of our intuitions about optimization depend on the
‘‘law of diminishing returns’’: the more of a resource ex-
Three Questions about the Model’s Theoretical Foundations
pended in a given effort, the less that each additional incre-
ment of allocation is able to accomplish. Very generally,optimization procedures converge because of diminishing re-
The S-K model focuses on differential allocation of energy
turns; in exceptional cases when diminishing returns fails,
between reproduction and somatic maintenance; however, as
the extremum is often obtained for variables at one end of
progressive caloric restriction is imposed, the mouse’s prin-
cipal adaptation is in neither of these variables, but rather in
Under conditions characterized by diminishing returns, a
body size. As total caloric intake is varied over a factor of
reduction in the total amount of any resource always results
three, body weight responds very nearly in proportion, and
in a reduction in each of its optimized apportionments. Spe-
it is not just adipose tissue that is affected, but muscle mass
cifically, we expect that an increasing food supply should
and most major organs, with notable exceptions of the brain
result in parallel increases in energy allocations for fertility
and the gonads (Weindruch and Sohal 1997). The net result
and for longevity. In two variants of the S-K model, an in-
is that the range of specific energy consumption (calories per
crease in total caloric budget results in a decrease of the
day per gram of lean body mass) varies by only 20%.
optimized portion allocated to somatic maintenance. Hidden
Life history analysis can tell us how longevity and fertility
in their assumptions is a violation of diminishing returns, in
are related to fitness; but the relevance of size is much less
the form of an effective fertility that varies quadratically with
clear and, presumably, much less direct. The challenge not
available caloric energy. Since the success of the model de-
addressed by the S-K model may be stated: if energy allo-
pends so critically on this relationship, the point becomes
cation is optimized under genetic control to maximize fitness,
crucial: why should each incremental increase in ambient
then why, as feeding makes more energy available, is such
a large proportion invested neither in fertility nor longevity
food intake corresponds to more calories available for repair
and maintenance. Only in the context of the model’s leastcredible variant is the observed sign for the relationship be-
Is the measure of fitness appropriate?
tween food intake and longevity correctly reproduced; andeven so, the result applies only to lactating females and only
Consistent with common usage in the field of life histories,
within a narrow range of the caloric intake variable. The DS
the authors have applied a definition of fitness equivalent to
theory remains an appealing and intuitively reasonable hy-
the Malthusian parameter r in the Euler-Lotka equation, as
pothesis; however, it may not be easily reconciled with a
developed by Fisher (1958). It is appropriate to apply r-se-
broad body of experimental data on caloric restriction.
lection to exponentially expanding populations; it places apremium on early reproduction, because each offspring is
weighted by a factor eϪrt in its contribution to fitness. Theopposite extreme is K-selection, appropriate to steady-state
Austad, S. 1995. Aging and caloric restriction: human effects and
mode of action. Neurobiol. Aging, 16:851–852.
Benton, T. G. and A. Grant. 2000. Evolutionary fitness in ecology:
Despite the ubiquity of Euler-Lotka in the literature, the
comparing measures of fitness in stochastic, density-dependent
use of r in steady-state environments (where, by definition,
environments. Evol. Ecol. Res. 2:769–789. r is identically equal to zero) entails paradoxes that have
Bronson, F. H. 1989. Mammalian reproductive biology. Univ. of
never been resolved. In fact, K may be very generally a more
Charlesworth, B. 1994. The evolution of life histories. Cambridge
robust measure of fitness than r, both as a theoretical tool
and a model of real-world selection (Benton and Grant 2000).
De Paolo, L. V. 1993. Dietary modulation of reproductive function. K-selection makes any pleiotropic theory of senescence (in-
Pp 221–246 in B. P. Yu. Modulation of aging processes by
cluding DS) more difficult to support (Mitteldorf, unpubl.
dietary restriction, CRC Press, Cleveland, OH.
Else, P. L. and A. J. Hulbert. 1987. Evolution of mammalian en-
dothermic metabolism—leaky membranes as a source of heat. Am. J. Physiol. 253:1–7. Thermodynamics of temperature maintenance
Fisher, R. A. 1958. The genetical theory of natural selection. 2nd
One of the competing energy demands invoked by Shanley
Holliday, R. 1989. Food, reproduction and longevity: is the ex-
and Kirkwood is the generation of heat, protecting the animal
tended lifespan of calorie-restricted animals an evolutionary ad-aptation? BioEssays 10:125–127.
in winter from hypothermia. They cite evidence (Else and
Kirkwood, T. B. L. 1977. Evolution of ageing. Nature 270:301–304.
Hulbert 1987) that mammals deploy chemical energy reserves
Masoro, E. and S. Austad. 1996. The evolution of the antiaging
to maintain body temperature in the cold of winter. But low-
action of dietary restriction: a hypothesis. J. Gerontol. A 51:
grade heat should not be reckoned in the same accounting
with other metabolic energy demands. In fact, conversion of
McCarter, R. J. M. 1993. Effects of exercise and dietary restriction
on energy metabolism and longevity. Pp. 157–174 in B. P. Yu.
chemical energy to any other purpose generates low-grade
Modulation of aging processes by dietary restriction. CRC Press,
heat as a by-product, and does so with 100% efficiency. The
thermodynamic principle is the same as for a 500-watt vac-
Merry, B. J. and A. M. Holehan. 1981. Serum profiles of LH, FSH,
uum cleaner, which delivers to the environment the entire
testosterone and 5-alpha-DHT from 21 to 1000 days of age inad libitum fed and dietary restricted rats. Exp. Gerontol. 16:
500 watts as ambient heat, while cleaning the rug ‘‘for free.’’
The body’s need for temperature homeostasis is not a separate
Millar, J. S. 1987. Energy reserves in breeding small mammals. Pp.
demand on caloric energy, but could be filled, for example,
231–240 in A. S. I. Loudon and P. A. Racey, eds. Reproductive
by the waste heat from vigorous efforts in DNA repair, tissue
energetics in mammals. Clarendon Press, Oxford, U.K.
rebuilding, and free radical scavenging. That useless fuel
Ross, M. H. and G. Bras. 1975. Food preference and length of life.
burning has evolved instead is a fact that commands expla-
Shanley, D. P., and T. B. L. Kirkwood. 2000. Calorie restriction
nation; and on its face, this squandering of free energy casts
and aging: a life history analysis. Evolution 54:740–750.
a shadow on any hypothesis that the caloric metabolism has
Venkatraman, J. and G. Fernandes. 1993. Modulation of immune
function during aging by dietary restriction. Pp. 193–220 in B. P. Yu. Modulation of aging processes by dietary restriction. CRCPress, Cleveland, OH.
Weindruch, R. and R. S. Sohal. 1997. Calorie intake and aging. N.
The Shanley-Kirkwood model is a serious and careful ef-
Weindruch, R., R. L. Walford, S. Fligiel, and D. Guthrie. 1986. The
fort to explain the observed relationship between caloric in-
retardation of aging in mice by dietary restriction—longevity,
take and lifespan in the context of the Disposable Soma the-
cancer, immunity and lifetime energy intake. J. Nutr. 116:
ory. Nevertheless, most versions of the model stubbornly
predict what our intuition tells us is reasonable: that more
Jon Geir Høyersten Sinnslidelsenes historie – noen temaer og tablåer Psykiatriens historie er en broket historie med mange beretninger om galskap og melankoli. Folkelige tradisjoner for forståelse og hjelp har gått parallelt med en vitenskapelig tilnærming. Det skyld- og skambelagte omkring psykiske lidelser har gått igjen. Fokuseringen på psykiatrien som en eneste lang histor
Coronary CT Angiography Your physician has recommended that you undergo a pain-free, non-invasive, non-surgical procedure called CT Angiography at Insight Medical Imaging. Through the use of x-rays and computers, the CT scan is able to generate cross-sectional images of your heart and coronary arteries. These very detailed and accurate images can help physicians diagnose and treat disea
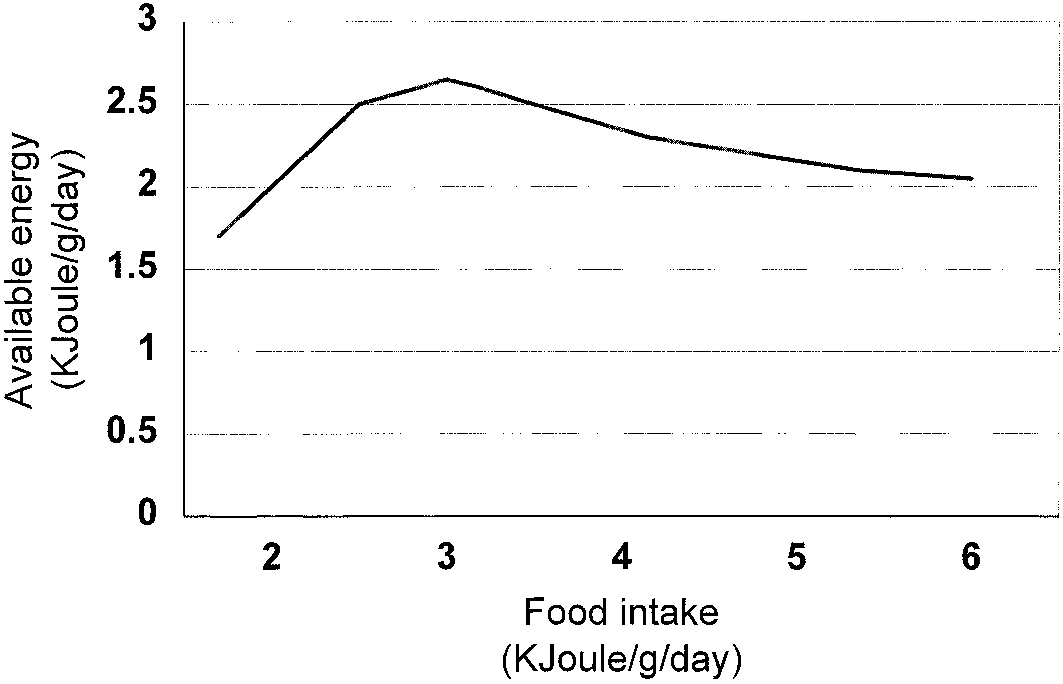 Life span of nonbreeding female mice increases smoothly
over a broad range as calories are reduced. Note that the increase
Results of the best-fitting version of the Shanley and Kirk-
continues substantially below the restriction level (80 K-cal) at
wood model. Energy that is not allocated to reproduction is con-
which female fertility is zero. This is difficult to reconcile with any
sidered available for somatic maintenance. Graph is for reproducing
model in which life-span extension derives from an energy reservoir
females. Note that the plot should not be directly compared to Figure
which is freed up as the animal diverts resources that had been
1 for this reason, and because both axes here have been normalized
devoted to reproduction. Graph adapted from Weindruch and Sohal
to body mass, which also varies with food intake. Graph adapted
1997, based on data from Weindruch et al. 1986.
Life span of nonbreeding female mice increases smoothly
over a broad range as calories are reduced. Note that the increase
Results of the best-fitting version of the Shanley and Kirk-
continues substantially below the restriction level (80 K-cal) at
wood model. Energy that is not allocated to reproduction is con-
which female fertility is zero. This is difficult to reconcile with any
sidered available for somatic maintenance. Graph is for reproducing
model in which life-span extension derives from an energy reservoir
females. Note that the plot should not be directly compared to Figure
which is freed up as the animal diverts resources that had been
1 for this reason, and because both axes here have been normalized
devoted to reproduction. Graph adapted from Weindruch and Sohal
to body mass, which also varies with food intake. Graph adapted
1997, based on data from Weindruch et al. 1986.