La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Geology-biology.eu
Evolution of Character Displacement in Darwin's Finches The following resources related to this article are available online at www.sciencemag.org (this information is current as of February 3, 2008 ):
including high-resolution figures, can be found in the online
Updated information and services, version of this article at: Supporting Online Material
A list of selected additional articles on the Science Web sites related to this article can be found at:
cites 25 articles
24 article(s) on the ISI Web of Science.
4 articles hosted by HighWire Press; see:
subject collections reprints
of this article or about obtaining permission to reproduce this article in whole or in part can be found at:
(print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by the
ScienceAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. Copyright2006 by the American Association for the Advancement of Science; all rights reserved. The title
have anything but a mild competitive effect onG. fortis. Their numbers gradually increased asa result of local production of recruits, aug-
mented by additional immigrants (22, 25), andreached a maximum of 354 T 47 (SE) in 2003(Fig. 3). Little rain fell in 2003 (16 mm) and
2004 (25 mm), there was no breeding in eitheryear, numbers of both species declined drasti-
Competitor species can have evolutionary effects on each other that result in ecological
cally, and from 2004 to 2005 G. fortis ex-
character displacement; that is, divergence in resource-exploiting traits such as jaws and beaks.
perienced strong directional selection against
Nevertheless, the process of character displacement occurring in nature, from the initial encounter
of competitors to the evolutionary change in one or more of them, has not previously been
Selection differentials in G. fortis were uni-
investigated. Here we report that a Darwin’s finch species (Geospiza fortis) on an undisturbed
formly negative for both males and females
Gala´pagos island diverged in beak size from a competitor species (G. magnirostris) 22 years after
treated separately (Table 2). Average selection
the competitor’s arrival, when they jointly and severely depleted the food supply. The observed
differentials in standard deviation units for the
evolutionary response to natural selection was the strongest recorded in 33 years of study, and
six measured traits that quantify bill size and
close to the value predicted from the high heritability of beak size. These findings support the role
shape and body size were 0.774 for males and
of competition in models of community assembly, speciation, and adaptive radiations.
0.649 for females. Compared with values re-ported in other studies elsewhere (27), they are
Characterdisplacement(1,2)isanevolu- termined by the tradeoff in energetic rewards unusually large. The six traits are positively
tionary divergence in resource-exploiting
from feeding on small and large seeds, and the
correlated to varying degrees. Selection gradi-
tradeoff is affected by variation in beak mor-
ent analysis helps to identify which particular
caused by interspecific competition (3–5). It has
traits were subject to selection independent of
the potential to explain nonrandom patterns of
replenishment (7, 18, 19). Competitors can mod-
correlations among traits (28). However, bill
co-occurrence and morphological differences
depth and width are so strongly correlated in
between coexisting species (6–10). Supporting
these samples (r 0 0.861 for males, 0.946 for
evidence has come from phylogenetic analyses
with the arrival of a new competitor species,
females) that their independent effects on sur-
(11) and from experimental studies of stickle-
setting up the potential for character displace-
vival cannot be distinguished. Selection gradient
backs, in which the role of directional selection
ment to occur. Between 1973 and 1982, a few
analysis without these two variables shows bill
in character divergence has been demonstrated
individuals of the large ground finch (G.
length to be the only significant entry into the
(12). The process of character displacement
magnirostris; È30 g) visited the island for
gradient, for both males Epartial regression co-
occurring in nature, from the initial encounter
short periods in the dry season but never bred
efficient (b) 0 –0.931 T 0.334 SE, P 0 0.0079;
of competitors to the evolutionary change in
(15). In late 1982, a breeding population was
R2 0 0.190^ and females (b 0 –0.814 T 0.295, P 0
one or more of them as a result of direction-
established by two females and three males at
al natural selection, has not previously been
the beginning of an exceptionally strong El
NiDo event that brought abundant rain to the
Table 1. Proportions of seeds in the diets of three
The situation on the small Gal"pagos island
island (1359 mm) (20–22). G. magnirostris is
finch species. Small seeds are a composite group
of Daphne Major (0.34 km2) has been referred
a potential competitor as a result of diet
of 22 species, medium seeds are O. echios, and
to as the classical case of character release
overlap with G. fortis (Table 1), especially in
large seeds are T. cistoides. N is the number of
(1, 2, 13), which is the converse of character
the dry season when food supply is limiting
observations. There is strong heterogeneity in the
displacement. Here, in the virtual absence of
(9, 23). The principal food of G. magnirostris
the small ground finch (Geospiza fuliginosa;
is the seeds of Tribulus cistoides, contained
0.0001). The reduction in G. fortis feeding on
weighing È12 g) and released from compe-
within a hard mericarp and exposed when a
Tribulus in 2004 makes a significant contribution
tition, the medium ground finch (G. fortis;
finch cracks or tears away the woody outer
(X 2 0 3.912, P G 0.05). Data were obtained by
È18 g) is unusually small in beak and body size.
covering (Fig. 1). Large-beaked members of
observations in the first 3 months of each year. In 1977 (only), when G. fortis experienced direc-
Lack (14) proposed that its small size reflects
the G. fortis population are capable of this
tional selection against small bill size, the pro-
an evolutionary shift enabling G. fortis to take
maneuver—indeed, survival in the 1977 drought
portion of large seeds in the diet rose to 0.304
to a large extent depended on it (13, 16)—but
available by the absence of its competitor. Sub-
on average they take three times longer than G.
sequent field studies demonstrated an associa-
magnirostris to gain a seed reward (13, 24).
tion, previously only inferred, between beak
The smallest G. fortis never attempt to crack
sizes and seed diets (13, 15). In 1977, a drought
them (18, 24). G. magnirostris compete with G.
on Daphne revealed that small seeds are pre-
ferred when they are abundant, but when they
Tribulus feeding sites and by reducing the
are scarce, finches turn increasingly to large
density of Tribulus fruits to the point at which
and hard seeds that only the large-beaked mem-
it is not profitable for G. fortis to feed on them,
bers of the population can crack (13, 15). Most
owing to handling inefficiencies in relation to
finches died that year, and mortality was heavi-
search and metabolic costs (7, 13, 18, 24). By
est among those with small beaks (13, 16, 17).
depleting the supply of Tribulus fruits, G.
Thus, a population_s mean beak size is de-
magnirostris was predicted to cause a selective
shift in G. fortis in the direction of small beak
Department of Ecology and Evolutionary Biology, Princeton
University, Princeton, NJ 08544–1003, USA.
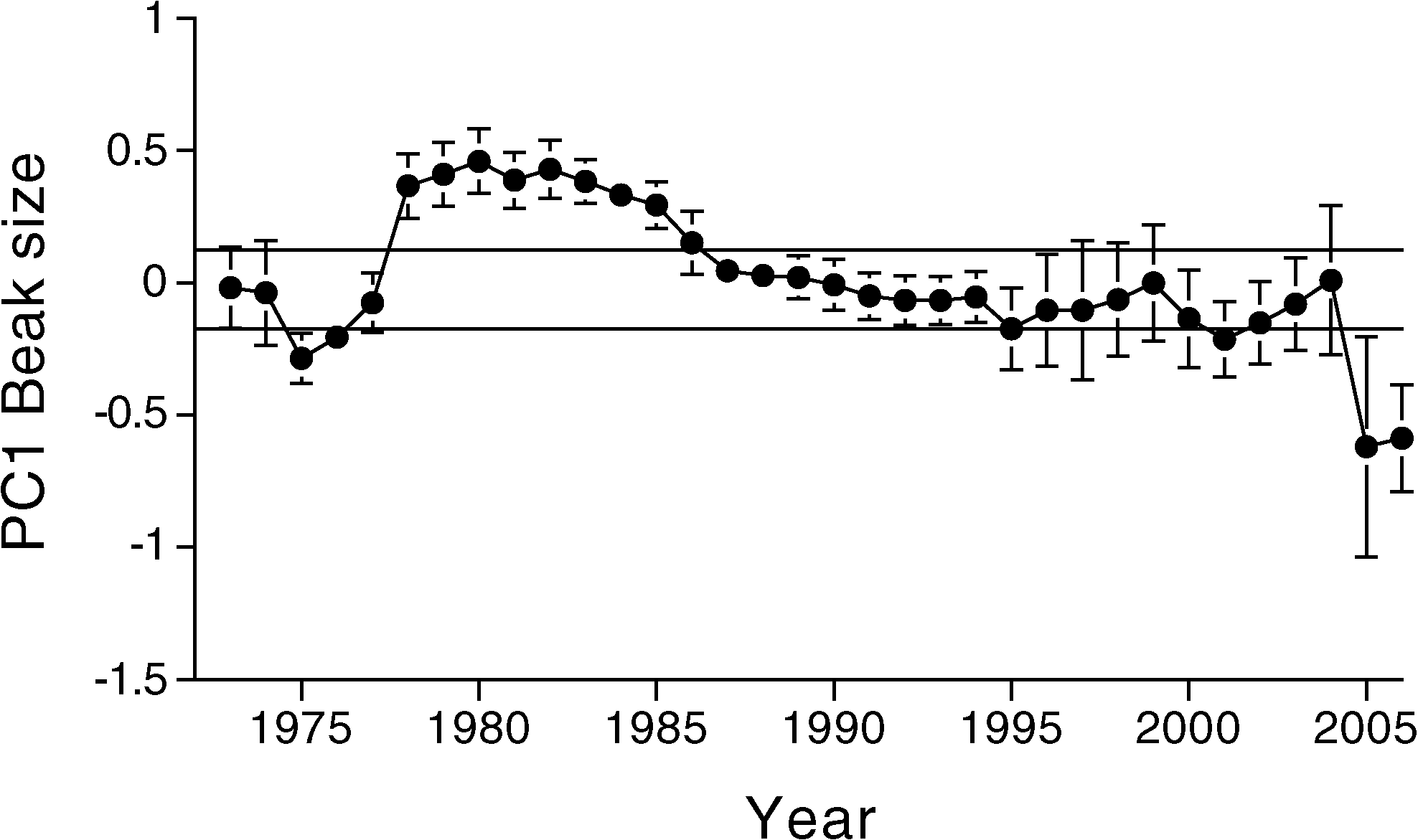
The predicted shift occurred in 2004 (Fig. 2).
Initially, the population size of G. magnirostris
*To whom correspondence should be addressed: E-mail:
was too small in relation to the food supply to
0.0130; R2 0 0.455). Inclusion of either bill
Thus, character displacement in G. fortis
consumed by a G. magnirostris individual each
depth or bill width made no difference to
occurred in 2004–2005. Four lines of evidence
day are sufficient for two G. fortis individuals if
these results. Overall bill size rather than bill
support the causal role of G. magnirostris. First,
they feed on nothing else (13). Moreover, a
length is identified as the most important fac-
the potential impact of G. magnirostris was
much higher fraction of G. magnirostris than G.
tor distinguishing survivors from nonsurvi-
greatest at the beginning of 2004 because their
fortis feed on Tribulus, as inferred from feeding
numbers (150 T 19) were closer to those of G.
observations (Table 1). As a result of their joint
size) was a selected trait in both sexes, where-
fortis (235 T 46) than at any other time (Fig. 3),
reduction of seed biomass, G. fortis fed on
and their population biomass was about the
Tribulus in 2004 only half as frequently as in
either. There was little effect on body size,
same, because a G. magnirostris individual was
other years (Table 1). We did not quantify food
unlike in the 1977 episode. In contrast to G.
approximately twice the mass of a G. fortis
supply; nevertheless, food scarcity was evident
fortis, the heavy mortality experienced by G.
from the exceptionally low feeding rates of G.
magnirostris was apparently not selective: Four
Second, G. magnirostris are largely depen-
magnirostris. In 2004, a minimum of 90 in-
surviving males did not differ from 32 non-
dent on an important food resource, Tribulus
dividuals were observed foraging for Tribulus
survivors in any of the six measured traits (all
seeds, which are not renewed during droughts.
mericarps for 200 to 300 s, and none obtained
P 9 0.1), and only 1 of 38 measured females
G. magnirostris deplete the Tribulus seed sup-
seeds from more than two mericarps; whereas
ply faster than do G. fortis. The seeds that are
under the more typical conditions prevailing inthe 1970s, a total of eight birds observed for thesame length of time fed on 9 to 22 mericarps,with an average interval between successivemericarps of only 5.5 T 0.5 s (SE) (24).
Third, numbers of G. fortis declined to a
lower level (83) in 2005 than at any time sincethe study began in 1973, and numbers of G. magnirostris declined so strongly from the2003 maximum that by 2005, only fourfemales and nine males were left. The popu-lation was almost extinct, apparently as a
result of exhaustion of the standing crop oflarge seeds and subsequent starvation. Of the137 G. magnirostris that disappeared in 2004–2005, 13.0% were found dead, and so were21.7% of 152 G. fortis. Consistent with thestarvation hypothesis, the stomachs of all deadbirds (23 G. magnirostris and 45 G. fortis,banded and not banded individuals combined)were empty.
The principal alternative food for both spe-
cies is the seeds of Opuntia cactus, butproduction in 2004 was low, the fourth lowestsince records were first kept systematically in1982 (23). Not only were cactus seeds insuffi-cient for the two granivore species to escape the
Fig. 1. Large-beaked G. fortis (A) and G. magnirostris (B) can crack or tear the woody tissues of
dilemma of a diminishing supply of their pre-
T. cistoides mericarps (D), whereas small-beaked G. fortis (C) cannot. Five mericarps constitute a
ferred foods, they were insufficient for the
single fruit. In (D), the left-hand mericarp is intact. The right-hand mericarp, viewed from the other
cactus specialist G. scandens (È20 g), whose
(mesial) side, has been exploited by a finch, exposing five locules from which seeds have been
numbers, like those of G. fortis, fell lower (to
extracted. Mericarps are È8 mm long and are shown at twice the magnification of the finches.
50) than in any of the preceding 32 years. The
only escape was available to the smallest, mostG. fuliginosa–like, members of the G. fortispopulation, which are known to feed like G.
fuliginosa on small seeds with little individual
energy reward (13, 18). We have no feeding
observations to indicate that they survived as a
result of feeding on the typical components of
the G. fuliginosa diet: the very small seeds of
(13, 15, 23). Nevertheless, it may be signifi-
cant that two G. fuliginosa individuals were
present on the island in 2004 and both survived
The fourth line of evidence is the contrast
sequent changes in themean. Sample sizes vary
between the directions of strong selection on
the G. fortis population in the presence (2004)
and near absence (1977) of G. magnirostris. In
values are reversed so that mean size increases from the origin.
1977, a year of only 24 mm of rain and no
breeding, body size and beak size of both male
be expected from strong directional selection
to null or neutral models (6, 9). Replicated
and female G. fortis considered separately were
against large size (32). This was observed. The
subject to selection (Table 2). Average selec-
mean beak size (PC1 ) of the 2005 generation
needed to demonstrate definitively the causal
tion differentials were 0.642 for males and
measured in 2006 was significantly smaller
role of competition, not only as an ingredient
0.668 for females, and they were uniformly
than that in the 2004 sample of the parental
of natural selection of resource-exploiting
positive. In the intervening years, 1978–2003,
traits (12) but as a factor in their evolution
there was a weaker selection episode favoring
0.0001). The difference between generations is
(33). Our findings should prove useful in
0.70 SD, which is exceptionally large (27, 29).
designing realistic experiments, by identifying
It may be compared with the range of values
ecological context (high densities at the start of an
small seeds and scarcity of large ones after the
predicted from the breeders equation, namely
environmental stress) and by estimating the
El NiDo event of 1982–1983 (20, 21, 23). At
the product of the average selection differential
that time, G. magnirostris were rare (22, 25);
numbers varied from 2 to 24. The selection
intervals of the heritability estimate. The ob-
events of 1977 and 2004 stand out against a
1. W. L. Brown Jr., E. O. Wilson, Syst. Zool. 5, 49
background of relative morphological stability
predicted range of 0.66 to 1.00 SD. Although
(29) (Fig. 2). Immediately before 2004 there
a small component of the response is probably
2. P. R. Grant, Biol. J. Linn. Soc. 4, 39 (1972).
was no unusual rainfall to cause a change in the
attributable to environmental factors Efood
3. B. W. Robinson, D. S. Wilson, Am. Nat. 144, 596
composition of the food supply and no other
supply and finch density (30, 32)^, the major
4. D. C. Adams, F. J. Rohlf, Proc. Natl. Acad. Sci. U.S.A. 97,
unusual environmental factor such as tempera-
component is genetic. This is the strongest
ture extremes or an invasion of predators, yet
evolutionary change seen in the 33 years of the
5. D. W. Pfennig, P. J. Murphy, Ecology 84, 1288
with the same amount of rain as in 1977, and
6. T. W. Schoener, in Ecological Communities: Conceptual
The evolutionary changes that we observed
Issues and the Evidence, D. R. Strong, L. G. Abele,
environment, large finches survived at a high
A. B. Thistle, Eds. (Princeton Univ. Press, Princeton, NJ,
Lack. Nevertheless, they provide direct support
frequency in 2004. The conspicuous difference
for his emphasis on the ecological adjustments
7. D. Schluter, T. Price, P. R. Grant, Science 227, 1056
between these years was the number of G.
that competitor species make to each other,
8. J. B. Losos, Proc. Natl. Acad. Sci. U.S.A. 97, 5693
magnirostris: 2 to 14 occasional visitors in
specifically in the final stages of speciation and
1977 (15) versus 150 T 19 residents at the
more generally in adaptive radiations (9–12, 14).
9. P. R. Grant, Ecology and Evolution of Darwin’s Finches
They also support models of ecological com-
(Princeton Univ. Press, Princeton, NJ, 1999).
Given the high heritability of beak size of
munity assembly that incorporate evolutionary
10. D. Schluter, Am. Nat. 156, S4 (2002). 11. J. B. Losos, Evolution 44, 588 (1990).
G. fortis (30, 31), an evolutionary response is to
effects of interspecific competition, in contrast
12. D. Schluter, Science 266, 798 (1994). 13. P. T. Boag, P. R. Grant, Biol. J. Linn. Soc. 22, 243
14. D. Lack, Darwin’s Finches (Cambridge Univ. Press,
15. P. T. Boag, P. R. Grant, Ecol. Monogr. 54, 463
16. P. T. Boag, P. R. Grant, Science 214, 82 (1981).
17. T. D. Price et al., Nature 309, 787 (1984).
18. T. Price, Ecology 68, 1015 (1987).
19. C. W. Benkman, Ecol. Monogr. 57, 251 (1987). 20. H. L. Gibbs, P. R. Grant, J. Anim. Ecol. 56, 797
21. H. L. Gibbs, P. R. Grant, Nature 327, 511 (1987).
22. P. R. Grant, B. R. Grant, Evolution 49, 229 (1995).
23. P. R. Grant, B. R. Grant, in Long-Term Studies of
Vertebrate Communities, M. L. Cody, J. A. Smallwood, Eds. (Academic Press, New York, 1996), pp. 343–390.
24. P. R. Grant, Anim. Behav. 29, 785 (1981). 25. P. R. Grant, B. R. Grant, K. Petren, Genetica 112-113,
Table 2. Selection differentials for G. fortis in the presence (2004) and absence (1977) of G.
26. See methods in supporting material on Science Online. 27. J. G. Kingsolver et al., Am. Nat. 157, 245 (2001).
magnirostris. Statistical significance at P G 0.05, G0.01, G0.005, and G0.001 is indicated by *, **,
28. R. Lande, S. Arnold, Evolution 37, 1210 (1983).
29. P. R. Grant, B. R. Grant, Science 296, 707 (2002).
30. P. T. Boag, Evolution 37, 877 (1983). 31. L. F. Keller et al., Heredity 87, 325 (2001).
32. P. R. Grant, B. R. Grant, Evolution 49, 241 (1995). 33. P. R. Grant, Science 266, 802 (1994).
34. We thank K. T. Grant, L. F. Keller, K. Petren, and U. Reyer
for help with recent fieldwork, and the Charles Darwin
Research Station and Gala´pagos National Park Service forpermission and support. The research was supported by
www.sciencemag.org/cgi/content/full/313/5784/224/DC1
5 April 2006; accepted 25 May 200610.1126/science.1128374
Properties and Performance of Rear Screen MaterialsStewart AeroGlas 100 vs. Da Lite DA 100 WAOne area of the audio visual industry that requires further study involves published specifications of products and how these specifications relate to the products as delivered and their actual performance in systems integration. The two areas of concern in our current studies involves video projectors a
www.epjournal.net – 2010. 8(2): 275-283 ¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯¯ Why it’s interesting why women have sex A review of Cindy M. Meston and David M. Buss, Why Women Have Sex: The Psychology of Sex in Women’s Own Voices. Times Books: New York, 2009, 306 pp.,US$25.00, ISBN 978-0-8050-8834-2 (hardcover). J. Brett Smith, Evolution Working Group, University of Ala

 Evolution of Character Displacement in Darwin's
Evolution of Character Displacement in Darwin's
 0.0130; R2 0 0.455). Inclusion of either bill
Thus, character displacement in G. fortis
consumed by a G. magnirostris individual each
depth or bill width made no difference to
occurred in 2004–2005. Four lines of evidence
day are sufficient for two G. fortis individuals if
these results. Overall bill size rather than bill
support the causal role of G. magnirostris. First,
they feed on nothing else (13). Moreover, a
length is identified as the most important fac-
the potential impact of G. magnirostris was
much higher fraction of G. magnirostris than G.
0.0130; R2 0 0.455). Inclusion of either bill
Thus, character displacement in G. fortis
consumed by a G. magnirostris individual each
depth or bill width made no difference to
occurred in 2004–2005. Four lines of evidence
day are sufficient for two G. fortis individuals if
these results. Overall bill size rather than bill
support the causal role of G. magnirostris. First,
they feed on nothing else (13). Moreover, a
length is identified as the most important fac-
the potential impact of G. magnirostris was
much higher fraction of G. magnirostris than G.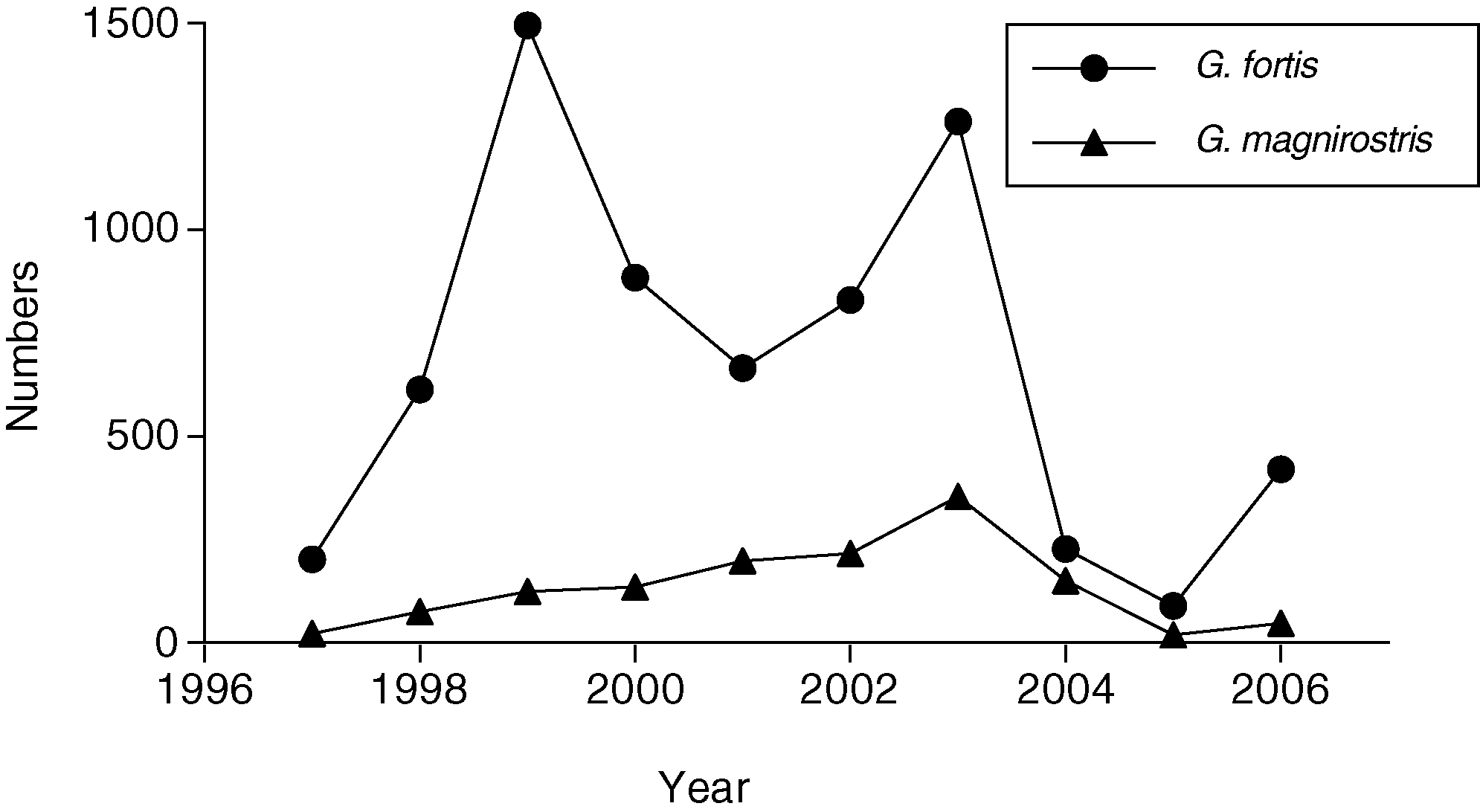 breeding, body size and beak size of both male
be expected from strong directional selection
to null or neutral models (6, 9). Replicated
and female G. fortis considered separately were
against large size (32). This was observed. The
subject to selection (Table 2). Average selec-
mean beak size (PC1 ) of the 2005 generation
needed to demonstrate definitively the causal
tion differentials were 0.642 for males and
measured in 2006 was significantly smaller
role of competition, not only as an ingredient
0.668 for females, and they were uniformly
than that in the 2004 sample of the parental
of natural selection of resource-exploiting
positive. In the intervening years, 1978–2003,
traits (12) but as a factor in their evolution
there was a weaker selection episode favoring
0.0001). The difference between generations is
(33). Our findings should prove useful in
0.70 SD, which is exceptionally large (27, 29).
breeding, body size and beak size of both male
be expected from strong directional selection
to null or neutral models (6, 9). Replicated
and female G. fortis considered separately were
against large size (32). This was observed. The
subject to selection (Table 2). Average selec-
mean beak size (PC1 ) of the 2005 generation
needed to demonstrate definitively the causal
tion differentials were 0.642 for males and
measured in 2006 was significantly smaller
role of competition, not only as an ingredient
0.668 for females, and they were uniformly
than that in the 2004 sample of the parental
of natural selection of resource-exploiting
positive. In the intervening years, 1978–2003,
traits (12) but as a factor in their evolution
there was a weaker selection episode favoring
0.0001). The difference between generations is
(33). Our findings should prove useful in
0.70 SD, which is exceptionally large (27, 29).