La tétracycline, connue sous le nom commercial Sumycin, agit en bloquant la fixation de l’ARNt sur la sous-unité 30S ribosomale, interrompant l’élongation de la chaîne protéique bactérienne. Ce mécanisme confère une activité sur un spectre large, incluant bactéries Gram positives, Gram négatives, rickettsies et spirochètes. Sa biodisponibilité digestive varie selon la prise alimentaire et les interactions avec les ions divalents comme calcium et magnésium. Sa diffusion tissulaire est importante, notamment dans les voies respiratoires et génito-urinaires. L’élimination se fait par voie rénale et biliaire. Les effets indésirables incluent photosensibilisation, troubles digestifs et coloration dentaire en cas d’administration précoce. Les guides thérapeutiques mentionnent sumycin prix, en soulignant la nécessité de restreindre son utilisation afin de limiter les résistances acquises.
Doi:10.1016/j.neulet.2004.02.040
Post-synaptic N-methyl-D-aspartate signalling in hippocampal neurons ofrat: spillover increases the impact of each spike in a short burst dischargeq
Sergei E. Grebenyuk, Natasha A. Lozovaya, Timur S. Tsintsadze, Oleg A. Krishtal*
Bogomoletz Institute of Physiology, 4, Bogomoletz St., 01024 Kiev, Ukraine
High-frequency burst discharges in hippocampus typically consist of less than ten spikes fired at frequencies too high to be followed by a
post-synaptic neuron. How significant are these numbers for synaptic signalling? We have measured the N-methyl-D-aspartate (NMDA)component of the excitatory post-synaptic current (EPSCNMDA) in hippocampal CA1 neurons of rat after burst discharge of variable duration. The synaptic facilitation is accompanied by a slow-down of the EPSCNMDA which develops on a spike-to-spike basis. Consequently thecharge transferred by the after-burst EPSCNMDA is increased with each spike. The phenomenon is most probably due to the spillover-mediated recruitment of extrasynaptic NMDA receptors. In terms of post-synaptic signalling it dramatically increases the impact of eachspike in a short burst discharge.
q 2004 Elsevier Ireland Ltd. All rights reserved.
Keywords: N-Methyl-D-aspartate receptors; Hippocampus; CA3/CA1 synapses; Burst discharge; Extrasynaptic receptors; Spillover
Natural spiking patterns in the brain are composed of
this question, we analyzed a-amino-3-hydroxy-5 methyl-
relatively short periods of high-frequency activity. From
isoxazole-4-proprionic acid (AMPA) and an N-methyl-D-
hippocampus through cortex, high-frequency burst dis-
aspartate (NMDA) component of the excitatory post-
charges are believed to be associated with information
synaptic currents (EPSCs) in CA1 pyramidal neurons
processing and memory consolidation. Specifically in
elicited by short trains of high-frequency stimulation of
hippocampus, they have two to nine action potentials fired
at frequencies up to 200 Hz . How significant is every
This study was carried out on 21-day-old Wistar rats
spike within this small number? This question seems to be
(WAG/GSto, Moscow, Russia). After rapid decapitation, rat
especially important since at high frequencies of firing the
brains were immediately transferred to a Petri dish with a
post-synaptic neuron cannot follow pre-synaptic burst
chilled (4 8C) solution of the following composition: 120
discharge with an equivalent number of spikes. To address
mM NaCl, 5 mM KCl, 26 mM NaHCO3, 2 mM MgCl2 and20 mM glucose. The solution was constantly bubbled with a
q Laudation in honour of Professor Zimmermann Great thanks to
95% O2/5% CO2 gas mixture to maintain pH 7.4. During the
Professor Zimmermann from the senior author, Oleg A. Krishtal: I cannot
pre-incubation and experiments, the slices (300 – 400 mM
find a better word than ‘visionary’ to describe the Editorial activities of
thick) were kept fully submerged in the extracellular
Professor Zimmermann. At least on two occasions his openness to non-
solution: 135 mM NaCl, 5 mM KCl, 26 mM NaHCO3, 1.5
paradigmal findings was crucial to the first publication on the ‘proton
receptor and the ATP receptor in sensory neurons’ O.A. Krishtal, V.I.
Pidoplichko, A “receptor” for protons in small neurons of trigeminal
bubbled with 95% O2/5% CO2) at 30 –31 8C. Picrotoxin
ganglia: possible role in nociception. Neurosci. Lett., 24 (1981) 243 – 246
(25 – 50 mM) was added to the extracellular solution during
O.A. Krishtal, S.M. Marchenko, V.I. Pidoplichko, Receptor for ATP in the
the experiments to suppress the inhibitory activity of
membrane of mammalian sensory neurones. Neurosci. Lett., 35 (1983) 41 –
45. Both papers dealt with unusual findings and this was a penetratingintuition of the Editor to say ‘Yes’. Subsequent history demonstrated that he
Standard whole-cell patch clamp techniques were used to
was right: the first paper resulted in the cloning of ASICs and in establishing
record EPSCs from CA1 pyramidal neurons in situ in
their role as nociceptors, while the second one contributed to the role of the
response to stimulation of the Schaffer collateral/commis-
P2X receptor family in sensory function.
sural pathway. To prevent the spread of electrical activity
* Corresponding author. Tel.: þ 38-044-2532466; fax: þ 38-044-
from area CA3, mini-slices were prepared by making a cut
E-mail address: krishtal@serv.biph.kiev.ua (O.A. Krishtal).
orthogonal to the stratum pyramidale and extending to the
0304-3940/03/$ - see front matter q 2004 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.neulet.2004.02.040
S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
mossy fibres layer. The intracellular solution for patchpipettes contained 100 mM CsF, 40 mM NaH2PO4, 10 mMHEPES-CsOH, and 10 mM Tris – Cl (pH 7.2). N-(2,6-Dimethyl-phenylcarbamoylmethyl)-triethylammonium bro-mide (QX-314) (2 – 3 mM) was routinely added to theintracellular solution to block voltage-gated sodium con-ductance. Patch pipettes were pulled from soft borosilicateglass on a two-stage horizontal puller. When fire-polishedand filled with the intracellular solution, they had aresistance of 2 – 3 MV. To visualize cell bodies of CA1pyramidal neurons, the stratum oriens and alveus wereremoved by a saline jet from a micropipette. Currents weredigitally sampled at 400 ms intervals by a 12-digit ADCboard, filtered at 3 kHz, and stored on a hard disk for furtheranalysis. Access resistance was monitored throughout theexperiments and ranged typically from 6 to 9 MV. When theaccess resistance changed by more than 25% during theexperiment, the data were discarded. To stimulate a Schaffercollateral/commissural pathway, a bipolar Ni/Cr electrodewas positioned on the surface of the slice. Current pulses(10 – 100 mA) of 0.1 – 1 ms duration were delivered throughthe isolated stimulator HG 203 (Hi-Med, London, UK) at0.066 – 0.2 Hz.
Sodium bicarbonate and CsF were obtained from Merck
Fig. 1. Facilitation of EPSC by short trains of stimulation is accompanied
(Darmstadt, Germany); 4-AP, lidocaine and picrotoxin were
by a selective slow-down of the NMDA component. (A) (Left)
purchased from RBI (Natick, MA); 6-nitro-7-sulphamoyl-
Pharmacologically isolated EPSCAMPA evoked by a single stimulus applied
benzo[ f ]quinoxalin-2,3-dione (NBQX) was obtained from
to Schaffer collaterals is superimposed with the EPSCAMPA evoked by a ten
Tocris Cookson (Bristol, UK). All other chemicals were
pulses long (200 Hz) train (n ¼ 7). Here and below: stimulation protocols
are schematically represented over the traces; the insets demonstratenormalized time courses of the currents. (Right) The same for EPSC
When measured after a short train of stimuli delivered at
(n ¼ 30). (B) The kinetics of EPSCNMDA evoked by a single stimulus does
high frequency, kinetics of the AMPA component of EPSC
not depend on the strength of a single stimulus (left), but becomes stimulus-
are the same as after a single stimulus, while the NMDA-
dependent after seven pulses at 200 Hz (right) (n ¼ 10). The stimulus
intensities varied by a factor of six. (C) (Left) The slowdown of EPSC
goes a dramatic slow-down The changes in the
is not determined solely by the current amplitudes: the traces from (B)elicited by the largest single stimulus (*) and by the smallest stimulus in the
NMDA kinetics were quantified by normalizing the
train (**) have equal amplitudes (superimposed, not normalized). The
charge transfer with the peak amplitude of the EPSCNMDA
kinetics of the post-train EPSCNMDA is evidently slower. (Right) Slow-
(measured as the mean over a 10 ms window around the
down of EPSCNMDA decay under high-frequency stimulation is clearly seen
peak). A larger charge transfer corresponds to slower decay
at positive voltage þ 20 mV (n ¼ 4), when the voltage-dependent
kinetics and vice versa. The normalized charge transfer of
conductance is inactivated and voltage-dependent Mg2þ block of NMDAchannels is removed. The small current elicited by a single pulse is
NMDA was 355 ^ 32% of the same parameter for
normalized (*) to the peak value of current elicited by seven pulses at 200
NMDA induced by a single stimulus (EPSCNMDA)
Hz. (D) Theoretical changes in the EPSCNMDA kinetics due to the change in
the current amplitude. The plot shows the dependence of charge transferred
on the EPSCNMDA amplitude measured in the soma
attributed to inadequate voltage clamp. A biophysical model
of CA1 pyramidal cells was adapted to ourexperimental conditions in order to estimate the extent to
strates the responses of the same amplitude obtained after a
which the quality of the voltage clamp affects the EPSC
single stimulus and a burst stimulation. Their kinetics are
decay. The density of synapses in the model of CA1 neuron
obviously different. Therefore, at least qualitatively, we can
was varied and the EPSCNMDA amplitude, as well as the
rely on the experimental data: it is not the increase in the
changes in EPSCNMDA kinetics, were analyzed.
amplitude of the current that solely determines the slow-
demonstrates the theoretical relationship between the
down of EPSCNMDA, though, for a given number of stimuli,
amplitude and kinetics of EPSCNMDA. The experimentally
the larger the stimulus applied, the greater this effect
obtained changes in EPSCNMDA amplitude (, 1.5-fold
increase) can be associated with , 5% changes in the
The behaviour of EPSC components after the train (
calculated charge transfer vs. 355% changes in the
remarkably resembles the findings previously made in
experimentally detected charge transfer. demon-
conditions of enhanced transmitter release: the AMPA
S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
component has unaltered kinetics, while the NMDA
NMDA receptor saturation by single quanta and
component slows down and acquires stimulus-dependent
provide a basis for synaptic integration.
kinetics This phenomenology is consistent with the
Thus, the synaptic facilitation by a short train of
spillout of Glu from the synaptic cleft resulting in a cross-
stimulation imitating the natural pattern of burst dis-
talk between neighbouring synapses and/or
charge leads to the increased impact of spillover-
activation of extrasynaptic NMDA receptors If this
activated NMDA receptors. For a given number of
picture is correct, the enhanced release of Glu should have
stimuli in the train, the higher the frequency used (
the following consequences: (i) the contribution of extra-
synaptic receptors should increase; (ii) these receptors
should ‘see’ considerably lower concentrations of Glu as
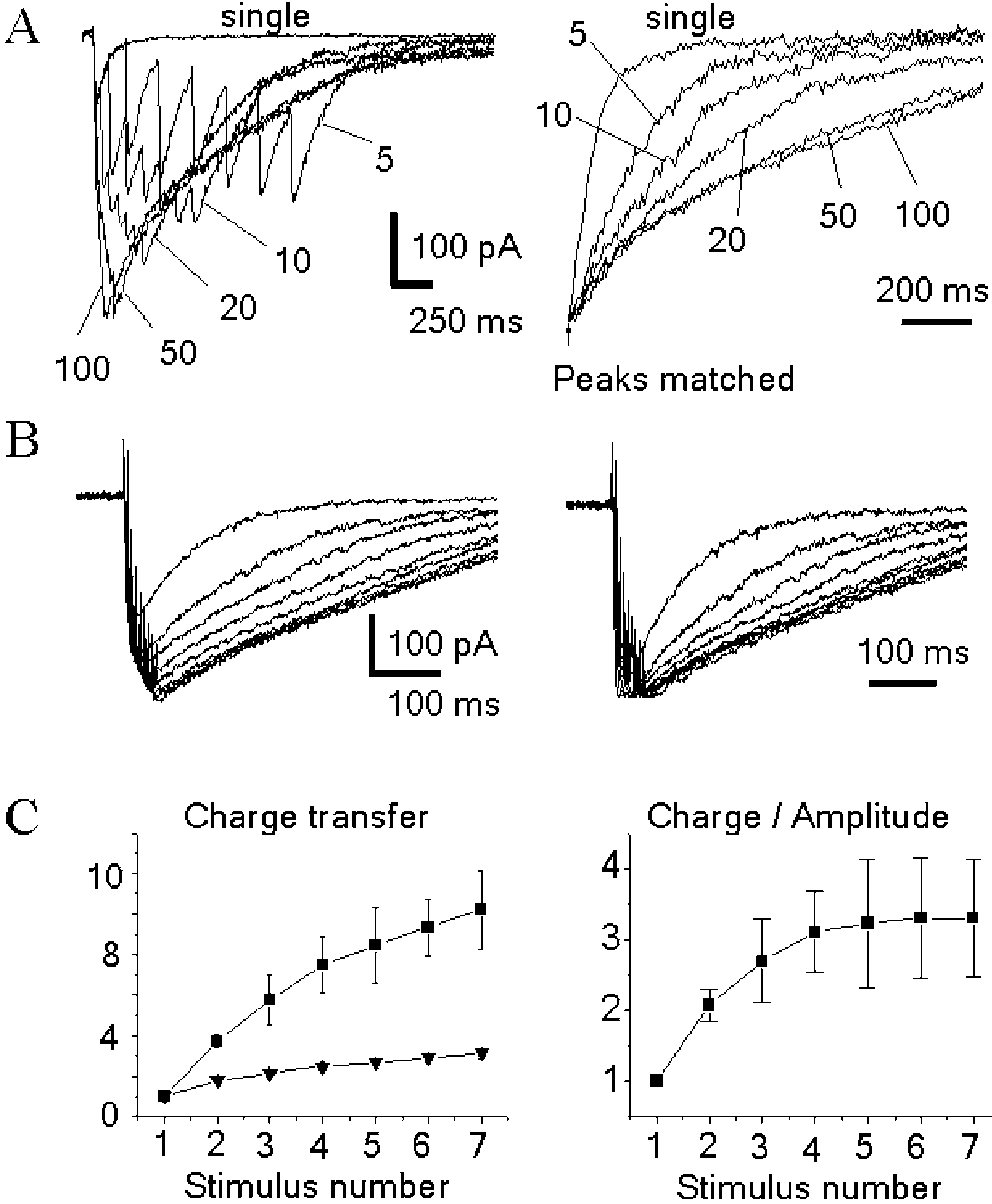
demonstrates the spike-to-spike changes in the
compared to the case of the receptors in the post-synaptic
EPSCNMDA. The kinetics becomes notably altered starting
already from the second stimulus. The charge transferred by
The efficacy of low-affinity competitive NMDA receptor
EPSCNMDA allows the estimation of the dependence of
antagonist D-amino adipate (D-AA) is known to depend on
Ca2þ entering the cell on the number of spikes in the pre-
the synaptic concentration of glutamate It has been used
synaptic burst discharge When compared with a
to distinguish receptors activated within active synapses
hypothetical dependence for the case of kinetics which
from those activated by spillover demonstrates
would remain unaltered by the burst discharge (only the
the experiments in which the prolonged after-train
EPSCNMDA was elicited in the presence of D-AA. The
steeper. Therefore, the information on the pre-synaptic
blocker leaves practically unaltered the kinetics of
activity (the number of spikes and of activated synapses) is
EPSCNMDA in the control, but markedly speeds up thedecay of EPSCNMDA induced by the train. Taken as ameasure of the change in the kinetics, normalized chargetransfer produced by the EPSCtrain
D-AA was 65 ^ 2% of control, while the correspondingvalue for the EPSCsingle
NMDA was 88 ^ 8% (P , 0:005, n ¼ 4).
Preferential inhibition of a slower component of theEPSCNMDA suggests that the NMDA receptors responsiblefor this component are activated by lower [Glu] than thosecontributing to the peak of the current.
We have found an indication of a dramatic increase in
the transmitter concentration acting at the peak of after-train EPSCNMDA. Inhibition of current by 50 mM D-AAin
(80 ^ 18%, n ¼ 4) at the peak of EPSCNMDA recordedafter a train of seven stimuli (The 1.25-folddecrease in EPSCNMDA amplitude observed in the D-AAexperiments should result in a 4% decrease in thetheoretical normalized charge transfer, whereas the actualchanges comprised 23%. The most straightforwardexplanation of this phenomenon is the induction of
multivesicular transmitter release in the course of high-
NMDA depends on the frequency of stimulation
and on the number of spikes in a train. (A) The slow-down of EPSCNMDA
frequency firing . Such a possibility can serve as
decay depends on the frequency of stimulation within the train. The
a functional ‘justification’ for the lack of synaptic
responses to seven stimuli at 5, 10, 20, 50 and 100 Hz are superimposed onthe left graph. Their normalized decays are on the right graph. (B) Theslow-down of EPSCNMDA on a spike-to-spike basis. (Left) Family ofEPSCNMDA evoked by the progressively increased number of pulses (one tonine) in the train (200 Hz). Holding voltage 2 45 mV. (Right)Corresponding traces are normalized and superimposed. (C) (Left) Thecharge transferred by after-burst EPSCNMDA depends on the number ofspikes (one to seven) in the train. The data obtained for five neuronsstimulated at 200 Hz (squares) are compared with the values obtained for
Fig. 2. D-AA accelerates after-burst EPSCNMDA. Block of EPSCNMDA by
the hypothetical case of unaltered kinetics (only amplitude facilitation is
D-AA induced by a single pulse (left) and by seven pulses at 200 Hz (right).
accounted for; triangles). (Right) The charge transferred by EPSCNMDA,
Holding voltage 2 45 mV. The insets demonstrate that D-AA does not alter
normalized to the peak amplitude. The dependence saturates after the 5th to
the kinetics of the current elicited by a single pulse, but speeds up the after-
6th stimulus. In the case of unaltered kinetics this dependence would
S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
synaptic transmission, in: C. Koch, I. Segev (Eds.), Methods in
(right) demonstrates that the gradual change in the
Neuronal Modeling, MIT Press, Cambridge, MA, 1998, pp. 1 – 25.
[6] J.S. Diamond, Neuronal glutamate transporters limit activation of
EPSCNMDA kinetics is saturated by the 5th to 6th stimulus.
NMDA receptors by neurotransmitter spillover on CA1 pyramidal
The experiments with Ca2þ imaging reveal that NMDA
cells, J. Neurosci. 21 (2001) 8328 – 8338.
receptors play a leading role in creating the post-synaptic
[7] E.R. Kandel, W.A. Spencer, Electrophysiology of hippocampal
Ca2þ signal and in controlling plasticity Spillover
neurons. II. Afterpotentials and repetitive firing, J. Neurophysiol. 24
seems to play an important role in shaping this signal by
[8] Y. Kovalchuk, J. Eilers, J. Lisman, A. Konnerth, NMDA receptor-
enhancing its dependence on the number of spikes in the
mediated subthreshold Ca(2 þ ) signals in spines of hippocampal
neurons, J. Neurosci. 20 (2000) 1791 – 1799.
[9] J.E. Lisman, Bursts as a unit of neural information: making unreliable
synapses reliable, Trends Neurosci. 20 (1997) 38 – 43.
[10] N.A. Lozovaya, M.V. Kopanitsa, Y.A. Boychuk, O.A. Krishtal,
Enhancement of glutamate release uncovers spillover-mediatedtransmission by N-methyl-D-aspartate receptors in the rat hippo-
This work was supported by the Wellcome Trust and
campus, Neuroscience 91 (1999) 1321 – 1330.
[11] Z.F. Mainen, R. Malinow, K. Svoboda, Synaptic calcium transients in
single spines indicate that NMDA receptors are not saturated, Nature399 (1999) 151 – 155.
[12] A.K. McAllister, C.F. Stevens, Nonsaturation of AMPA and NMDA
receptors at hippocampal synapses, Proc. Natl. Acad. Sci. USA 97(2000) 6173 – 6178.
[1] N. Arnth-Jensen, D. Jabaudon, M. Scanziani, Cooperation between
[13] M. Migliore, D.A. Hoffman, J.C. Magee, D. Johnston, Role of an A-
independent hippocampal synapses is controlled by glutamate uptake,
type K þ conductance in the back-propagation of action potentials in
Nat. Neurosci. 5 (2002) 325 – 331.
the dendrites of hippocampal pyramidal neurons, J. Comput.
[2] F. Asztely, G. Erdemli, D.M. Kullmann, Extrasynaptic glutamate
spillover in the hippocampus: dependence on temperature and the role
[14] T.G. Oertner, B.L. Sabatini, E.A. Nimchinsky, K. Svoboda,
of active glutamate uptake, Neuron 18 (1997) 281 – 293.
Facilitation at single synapses probed with optical quantal analysis,
[3] J.D. Clements, R.A. Lester, G. Tong, C.E. Jahr, G.L. Westbrook, The
Nat. Neurosci. 5 (2002) 657 – 664.
time course of glutamate in the synaptic cleft, Science 258 (1992)
[15] D.A. Rusakov, D.M. Kullmann, Extrasynaptic glutamate diffusion in
the hippocampus: ultrastructural constraints, uptake, and receptor
[4] R. Conti, J. Lisman, The high variance of AMPA receptor- and
activation, J. Neurosci. 18 (1998) 3158 – 3170.
NMDA receptor-mediated responses at single hippocampal synapses:
[16] S.S. Suzuki, G.K. Smith, Burst characteristics of hippocampal
evidence for multiquantal release, Proc. Natl. Acad. Sci. USA 100
complex spike cells in the awake rat, Exp. Neurol. 89 (1985) 90 – 95.
[17] G. Tong, C.E. Jahr, Multivesicular release from excitatory synapses of
[5] A. Destexhe, Z.F. Mainen, T.J. Sejnowski, Biophysical models of
cultured hippocampal neurons, Neuron 12 (1994) 51 – 59.
Getting It Up? Erectile Dysfunction & Lifestyle I have to tell you that in the past I have suffered from erectile dysfunction (ED)! It was AWFUL and resulted from the mental and emotional strain that emanated from financial problems (over three years’ worth of them). In a need to ‘escape reality’ I also ended up half-wrecking my body with alcohol and recreational drugs, which compl
Response, activates PLC->IP3,DAG=>Ca++ Similar to M1 mechanism. Sm. Muscles, secretory Agonists Action Bethanechol Stimulates GI and urinary bladder Carbachol + Increases ureteral peristalsis, contraction of detrusor muscle, and decreases Betanechol capacity of urinary bladder (increase in micturation) M3 agonist for salivary gland activation Induces marked salivation and diaphoresis (

 Post-synaptic N-methyl-D-aspartate signalling in hippocampal neurons ofrat: spillover increases the impact of each spike in a short burst dischargeq
Sergei E. Grebenyuk, Natasha A. Lozovaya, Timur S. Tsintsadze, Oleg A. Krishtal*
Bogomoletz Institute of Physiology, 4, Bogomoletz St., 01024 Kiev, Ukraine
High-frequency burst discharges in hippocampus typically consist of less than ten spikes fired at frequencies too high to be followed by a
post-synaptic neuron. How significant are these numbers for synaptic signalling? We have measured the N-methyl-D-aspartate (NMDA)component of the excitatory post-synaptic current (EPSCNMDA) in hippocampal CA1 neurons of rat after burst discharge of variable duration.
Post-synaptic N-methyl-D-aspartate signalling in hippocampal neurons ofrat: spillover increases the impact of each spike in a short burst dischargeq
Sergei E. Grebenyuk, Natasha A. Lozovaya, Timur S. Tsintsadze, Oleg A. Krishtal*
Bogomoletz Institute of Physiology, 4, Bogomoletz St., 01024 Kiev, Ukraine
High-frequency burst discharges in hippocampus typically consist of less than ten spikes fired at frequencies too high to be followed by a
post-synaptic neuron. How significant are these numbers for synaptic signalling? We have measured the N-methyl-D-aspartate (NMDA)component of the excitatory post-synaptic current (EPSCNMDA) in hippocampal CA1 neurons of rat after burst discharge of variable duration.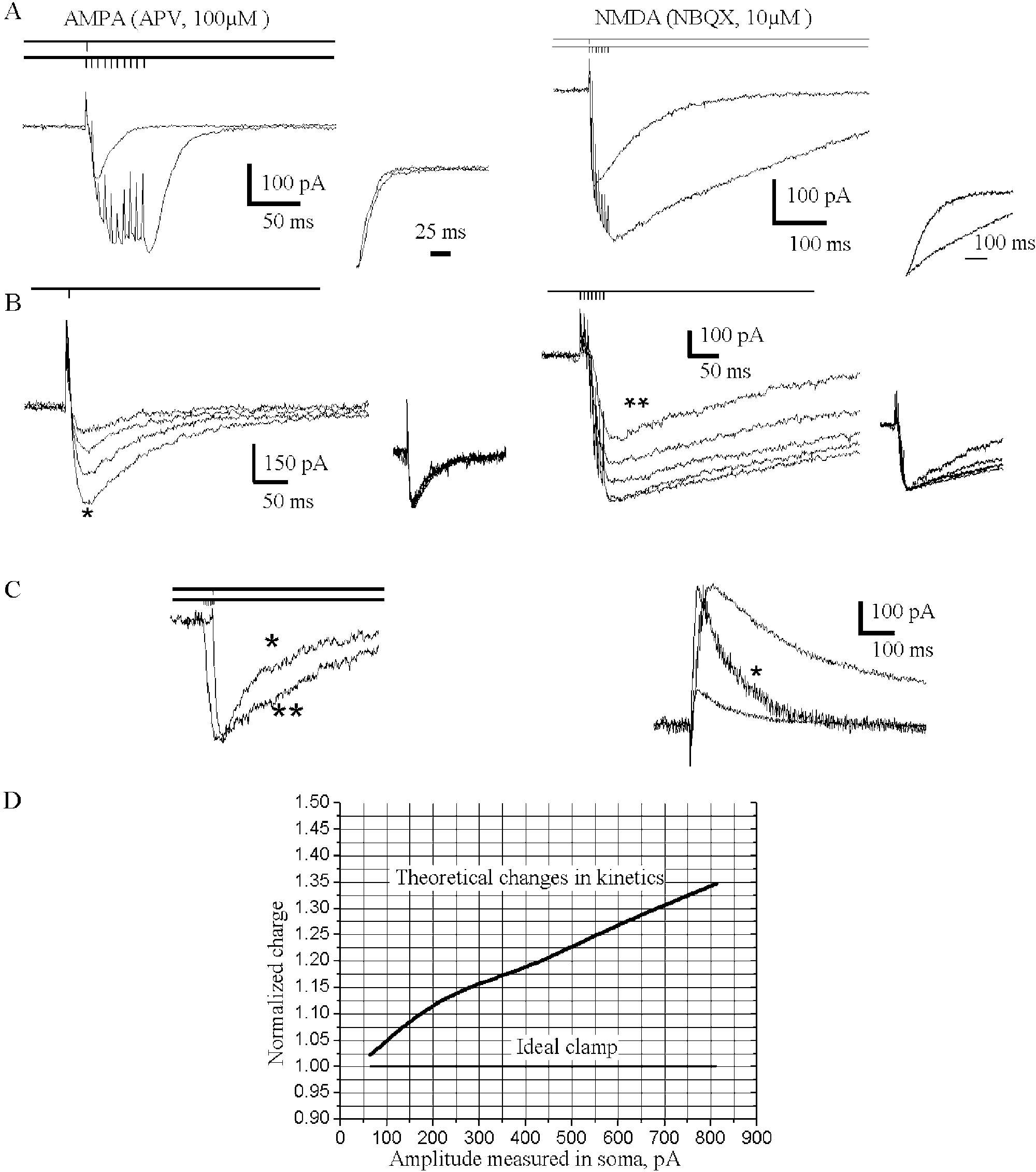 S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
mossy fibres layer. The intracellular solution for patchpipettes contained 100 mM CsF, 40 mM NaH2PO4, 10 mMHEPES-CsOH, and 10 mM Tris – Cl (pH 7.2). N-(2,6-Dimethyl-phenylcarbamoylmethyl)-triethylammonium bro-mide (QX-314) (2 – 3 mM) was routinely added to theintracellular solution to block voltage-gated sodium con-ductance. Patch pipettes were pulled from soft borosilicateglass on a two-stage horizontal puller. When fire-polishedand filled with the intracellular solution, they had aresistance of 2 – 3 MV. To visualize cell bodies of CA1pyramidal neurons, the stratum oriens and alveus wereremoved by a saline jet from a micropipette. Currents weredigitally sampled at 400 ms intervals by a 12-digit ADCboard, filtered at 3 kHz, and stored on a hard disk for furtheranalysis. Access resistance was monitored throughout theexperiments and ranged typically from 6 to 9 MV. When theaccess resistance changed by more than 25% during theexperiment, the data were discarded. To stimulate a Schaffercollateral/commissural pathway, a bipolar Ni/Cr electrodewas positioned on the surface of the slice. Current pulses(10 – 100 mA) of 0.1 – 1 ms duration were delivered throughthe isolated stimulator HG 203 (Hi-Med, London, UK) at0.066 – 0.2 Hz.
S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
mossy fibres layer. The intracellular solution for patchpipettes contained 100 mM CsF, 40 mM NaH2PO4, 10 mMHEPES-CsOH, and 10 mM Tris – Cl (pH 7.2). N-(2,6-Dimethyl-phenylcarbamoylmethyl)-triethylammonium bro-mide (QX-314) (2 – 3 mM) was routinely added to theintracellular solution to block voltage-gated sodium con-ductance. Patch pipettes were pulled from soft borosilicateglass on a two-stage horizontal puller. When fire-polishedand filled with the intracellular solution, they had aresistance of 2 – 3 MV. To visualize cell bodies of CA1pyramidal neurons, the stratum oriens and alveus wereremoved by a saline jet from a micropipette. Currents weredigitally sampled at 400 ms intervals by a 12-digit ADCboard, filtered at 3 kHz, and stored on a hard disk for furtheranalysis. Access resistance was monitored throughout theexperiments and ranged typically from 6 to 9 MV. When theaccess resistance changed by more than 25% during theexperiment, the data were discarded. To stimulate a Schaffercollateral/commissural pathway, a bipolar Ni/Cr electrodewas positioned on the surface of the slice. Current pulses(10 – 100 mA) of 0.1 – 1 ms duration were delivered throughthe isolated stimulator HG 203 (Hi-Med, London, UK) at0.066 – 0.2 Hz.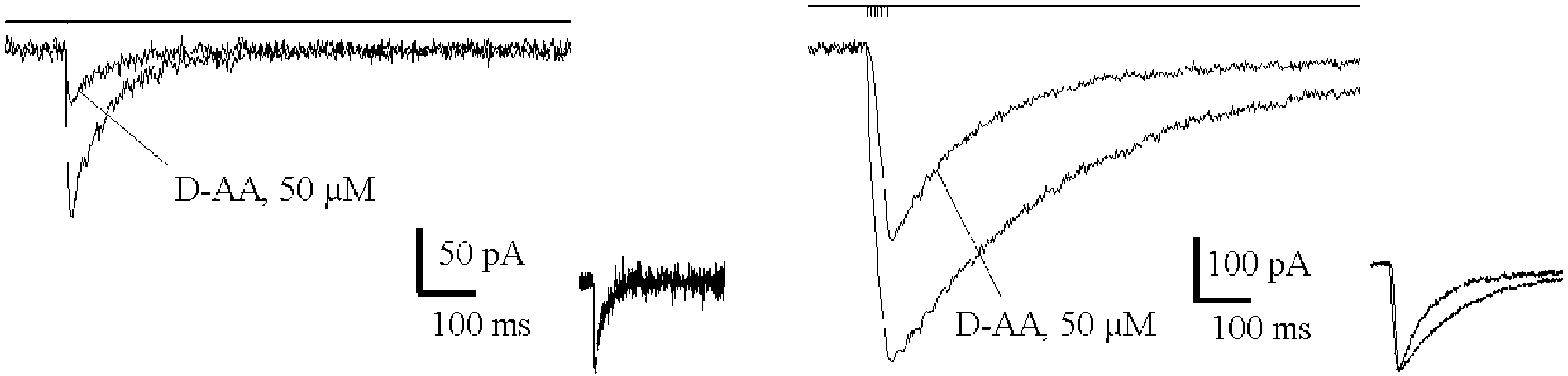
 S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
component has unaltered kinetics, while the NMDA
NMDA receptor saturation by single quanta and
component slows down and acquires stimulus-dependent
provide a basis for synaptic integration.
S.E. Grebenyuk et al. / Neuroscience Letters 361 (2004) 60–63
component has unaltered kinetics, while the NMDA
NMDA receptor saturation by single quanta and
component slows down and acquires stimulus-dependent
provide a basis for synaptic integration.